Погода в Городке на неделю (Минусинский район, Красноярский край)
В 02:30 в аэропорту «Абакан» (~34 км) было -28°C, замерзающий туман, дым, безветренно. Атм. давление на уровне аэропорта 755 мм рт.ст, влажность воздуха 75%.
В 01:00 на метеостанции «Минусинск» (~24 км) было -29°C, облачно, дымка, безветренно. Атмосферное давление на уровне станции 754 мм рт.ст, влажность воздуха 80%.
Сегодня в Городке ожидается −16°..−18°, облачно, без осадков, туман. Ночью −24°..−26°. Ветер восточный 1 м/с. Давление 761 мм рт.ст. Завтра −16°..−18°, значительная облачность, без осадков, туман. Ветер юго-восточный 1 м/с. Давление 762 мм рт.ст.
| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Утро | Утро−27° | пасмурнобез осадков | 100% | В, 1м/с | ||
| День | День−17° | облачнобез осадков, туман | 761 | 92% | ВЮВ, 1м/с | |
| Вечер−23° | облачно с прояснениямибез осадков | 762 | 100% | ВСВ, 1м/с |
| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь−25° | переменная облачностьбез осадков | 100% | ВЮВ, 1м/с | |||
| Утро | Утро−28° | облачно с прояснениямибез осадков | 100% | ВЮВ, 1м/с | ||
| День | День−17° | значительная облачностьбез осадков, туман | 762 | 90% | ЮВ, 1м/с | |
| Вечер | Вечер−23° | переменная облачностьбез осадков | 99% | ВСВ, 1м/с |
 ст.»>763
ст.»>763 ст.»>761
ст.»>761| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь | облачно с прояснениямибез осадков | 760 | 100% | ЮВ, 1м/с | ||
| Утро | Утро−20° | пасмурнобез осадков, туман | 98% | ВЮВ, 2м/с | ||
| День | День−12° | пасмурнобез осадков | 757 | 79% | ЮВ, 2м/с | |
| Вечер | Вечер−13° | пасмурнобез осадков, туман | 756 | ВЮВ, 2м/с |
 ст.»>759
ст.»>759| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь | Ночь−14° | пасмурнобез осадков | 95% | ВЮВ, 2м/с | ||
| День | День−10° | пасмурнобез осадков | 752 | 78% | ЮВ, 1м/с |

| Погода | Давл | Влж | Ветер | |||
|---|---|---|---|---|---|---|
| Ночь | Ночь−14° | пасмурнобез осадков, туман | 96% | ЮЮВ, 1м/с | ||
| День | День−10° | облачно с прояснениямибез осадков | 753 | 85% | СЗ, 2м/с |
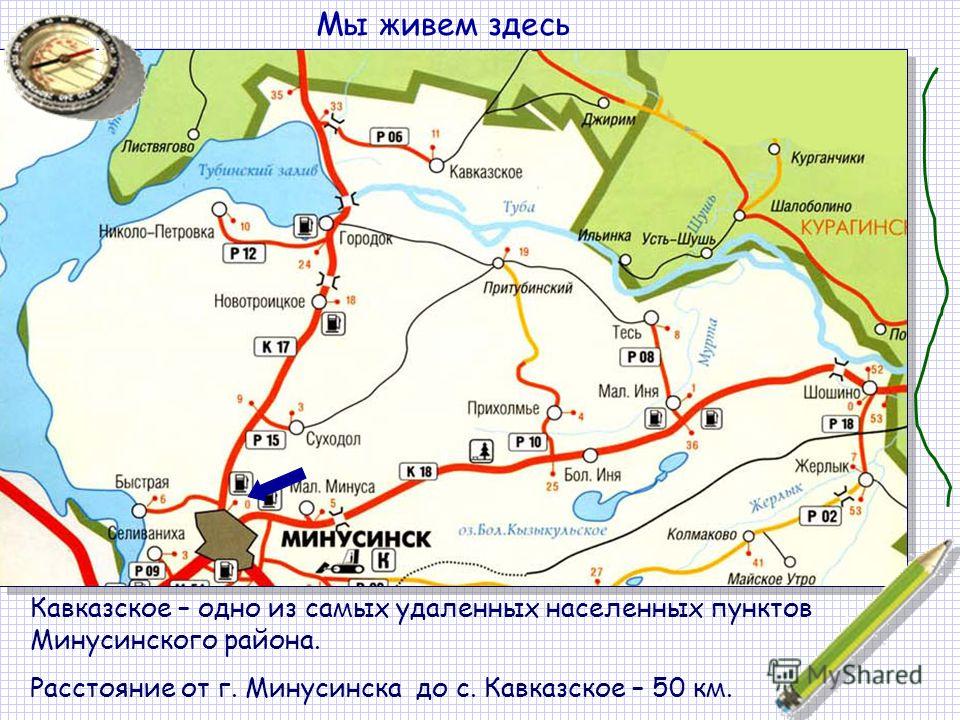 ст.»>748
ст.»>748| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь | Ночь−22° | переменная облачностьбез осадков | 100% | СВ, 1м/с | ||
| День | День−16° | пасмурнобез осадков | 756 | 95% | ВЮВ, 1м/с |
 ст.»>757
ст.»>757| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь | Ночь−25° | значительная облачностьбез осадков | 100% | В, 1м/с | ||
| День | День−16° | пасмурнобез осадков, туман | 752 | 94% | ВСВ, 1м/с |
 ст.»>755
ст.»>755| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь | Ночь−23° | пасмурнобез осадков | 100% | СВ, 1м/с | ||
| День | День−15° | пасмурнобез осадков | 750 | 91% | В, 1м/с |
 ст.»>751
ст.»>751| t°C | Погода | Давл | Влж | Ветер | ||
|---|---|---|---|---|---|---|
| Ночь | Ночь−17° | пасмурнонебольшой снег | 98% | ССВ, 1м/с |
 ст.»>749
ст.»>749Прогноз погоды в Городке на неделю (
- Красноярский край
- Минусинский район
- село Городок
| Температура | Осадки | Ветер | Влажность | Облачность | Давление | |
| 00:00 | -25,9° | сильная облачность | 0,7 м/с, Восток | 96% | 99% | 780 мм рт. ст. ст. |
| 01:00 | -25,3° | сильная облачность | 0,5 м/с, Восток | 96% | 98% | 780 мм рт. ст. |
| 02:00 | -24,6° | сильная облачность | 0,4 м/с, Восток | 96% | 98% | 780 мм рт. ст. |
| 03:00 | -23,2° | сильная облачность | 0,2 м/с, Восток | 96% | 98% | 780 мм рт. ст. |
| 04:00 | -23,3° | сильная облачность | 0,1 м/с, Восток | 93% | 97% | 780 мм рт. ст. |
| 05:00 | -23,4° | сильная облачность | 0,1 м/с, Восток | 90% | 97% | 779 мм рт. ст. |
| 06:00 | -23,5° | сильная облачность | 0,1 м/с, Восток | 84% | 97% | 778 мм рт. ст. |
| 07:00 | -22,8° | сильная облачность | 0,2 м/с, Северо-Восток | 84% | 95% | 778 мм рт. ст. ст. |
| 08:00 | -22,1° | сильная облачность | 0,2 м/с, Северо-Восток | 84% | 93% | 778 мм рт. ст. |
| 09:00 | -20,7° | сильная облачность | 0,4 м/с, Северо-Восток | 85% | 89% | 778 мм рт. ст. |
| 10:00 | -21,2° | сильная облачность | 0,5 м/с, Северо-Восток | 87% | 88% | 778 мм рт. ст. |
| 11:00 | -21,7° | сильная облачность | 0,6 м/с, Северо-Восток | 89% | 87% | 779 мм рт. ст. |
| 12:00 | -22,7° | сильная облачность | 0,7 м/с, Северо-Восток | 93% | 85% | 779 мм рт. ст. |
| 13:00 | -23,1° | сильная облачность | 0,6 м/с, Северо-Восток | 94% | 83% | 779 мм рт. ст. |
| 14:00 | -23,5° | сильная облачность | 0,4 м/с, Северо-Восток | 95% | 81% | 779 мм рт. ст. ст. |
| 15:00 | -24,3° | пасмурно | 0,2 м/с, Северо-Восток | 98% | 77% | 780 мм рт. ст. |
| 16:00 | -25,3° | пасмурно | 0,1 м/с, Восток | 94% | 80% | 780 мм рт. ст. |
| 17:00 | -26,2° | пасмурно | 0,1 м/с, Восток | 91% | 83% | 780 мм рт. ст. |
| 18:00 | -28,2° | сильная облачность | 0,0 м/с, Восток | 85% | 90% | 780 мм рт. ст. |
| 19:00 | -27,9° | сильная облачность | 0,0 м/с, Восток | 86% | 90% | 780 мм рт. ст. |
| 20:00 | -27,5° | сильная облачность | 0 м/с, Восток | 87% | 90% | 780 мм рт. ст. |
| 21:00 | -26,9° | сильная облачность | 0 м/с, Восток | 90% | 91% | 780 мм рт. ст. ст. |
| 22:00 | -26,8° | сильная облачность | 0,0 м/с, Восток | 91% | 91% | 781 мм рт. ст. |
| 23:00 | -26,7° | сильная облачность | 0,1 м/с, Восток | 92% | 91% | 781 мм рт. ст. |
Село Городок Минусинского района Красноярского края
Село Городок расположено в Минусинском районе Красноярского края.
Из архива panoramio . Права на фотографию принадлежат их владельцам
У Вас есть фото из Городка?
Добавьте их пожалуйста!
Краткая информация о Городке
Телефонный код: 39132
Автомобильный код: 24, 84, 88
Почтовый индекс: 662631
Код ОКАТО: 4233810000
Разница во времени с Москвой: +4
ч.
Фотографии Городка
Добавить фото Были в Городке и сделали снимки города и интересных мест? Разместите их! Многие посетители нашего сайта делают фото в своих поездках. Публикуйте свои фото из Городка и из других городов! Отмечайте если Вы тоже видели эти места! Теперь появилась возможность добавления с мобильной версии сайта, что намного облегчает перенос фото из со смартфонов на сайт!Сувениры из Городка
Добавить Сувенир Были в Городка и привезли сувениры? Покажите их всем! Магнитики на холодильник, тарелки, вымпелы, кружки и все что Вы привозите с собой из командировок и поездок в Городок! Размещайте и отмечайте если у вас уже есть такие ! Разместите их фото! С мобильной версии сайта это сделать намного проще и перенос фото магнитиков из Городка со смартфонов на сайт будет удобнее!Последние отзывы о Городке
Добавить отзыв Уважаемые посетители сайта Командировка. ру!
ру! Пока нет ни одного отзыва Напишите отзыв!
Последние вопросы о селе
Добавить вопросЕсли у Вас есть вопросы по данному населенному пункту или Вы ищете людей из селаГородка, проживающих в нем, Вы можете задать задать соответсвующий вопрос. Надеемся, что наши пользователи помогут Вам с ответами.
Енисейск, Сибирь (Правительство) — Энциклопедия
ЕНИСЕЙСК, губерния Восточной Сибири, простирающаяся от границы с Китаем до берегов Северного Ледовитого океана, площадью 986 908 кв.м. Имеет Тобольскую и Томскую губернии на З., Якутскую и Иркутскую на В., С.З. Монголия на Ю. и Северный Ледовитый океан на С. Ее южная оконечность, находящаяся на 51° 45′ с. ш., а северная (мыс Челюскин) на 77° 38′, сочетает в себе большое разнообразие орографических регионы в С. в тундры арктического побережья.
в тундры арктического побережья.
Бордюр высокого плато С.З. Монголия, известная под общим названием Западные Саяны и достигающая высот 7000—8000 футов, ограничивает его на Ю. Она окаймлена на северо-западном склоне зоной почти 100 м. широкий, характеризуется узкими долинами, разделяющими параллельные цепи гор, сложенных из кристаллических сланцев, высотой от 6000 до 7000 футов. Здесь в непроходимых лесах охотой живут несколько тунгусских семей.Южнее, в бассейнах рек Тубы, Сисима, Юса, Кана, Агула и Бирюсы, долины высокогорных массивов содержат богатые золотоносные месторождения, а вдоль тайги установлены многочисленные золотопромывки. Уплощенный горный хребет, едва достигающий высоты от 3000 до 35 00 футов, простирается на северо-восток. от Кузнецкого Алатау, и отделяет сухие степи Минусинска и Абакана от следующей террасы равнин высотой от 1200 до 1700 футов, которые также простираются на С.В. из Барнаула в Томской губернии до Красноярска и в верховья бассейна Вилюя.На внешней границе этой террасы, на пространстве между верхней Тунгузкой, или Ангарой, и Подкаменной Тунгузкой, возвышается другая система гор, известная как Енисейская тайга. Эта система состоит из нескольких параллельных цепочек, работающих на S.W. до северо-восточного, и достигают 2500-3500 футов над уровнем моря, хотя они намного ниже на левом берегу Енисея. Енисейская тайга на протяжении многих лет является одним из самых продуктивных золотоносных районов Сибири не столько по процентному содержанию золота в ее россыпных месторождениях (бедных по сравнению с Олекминскими), сколько по возможностям добычи золота. прииски с продовольствием, произведенным в степях Минусинска.
Эта система состоит из нескольких параллельных цепочек, работающих на S.W. до северо-восточного, и достигают 2500-3500 футов над уровнем моря, хотя они намного ниже на левом берегу Енисея. Енисейская тайга на протяжении многих лет является одним из самых продуктивных золотоносных районов Сибири не столько по процентному содержанию золота в ее россыпных месторождениях (бедных по сравнению с Олекминскими), сколько по возможностям добычи золота. прииски с продовольствием, произведенным в степях Минусинска.
За Енисейской тайгой начинаются низменности, нигде не возвышающиеся над уровнем моря более чем на несколько сотен футов. Они полого наклоняются к Северному Ледовитому океану и покрыты озерами, скудными лесами и болотами; и по мере приближения к океану они все более и более приобретают характер бесплодной тундры. За 70° с.ш. деревья встречаются только по руслам рек. Два хребта, однако, нарушают однообразие низменностей — Тунгусский, протянувшийся на С.-В., между реками Хатангой и Анабаром, и горы Бырранга, огибающие С. Западный берег полуострова Таймыр. Берега Северного Ледовитого океана изрезаны глубокими лиманами, в том числе Тазовского, проникающего на 600 м. в глубь материка, а Енисея на 300 м. Таймырский, Фаддейский и Хатангский заливы представляют собой широкие и глубокие котловины, почти круглый год скованные льдом. Полуостров Таймыр, между Енисеем и Хатангой, представляет собой каменистую тундру.
Западный берег полуострова Таймыр. Берега Северного Ледовитого океана изрезаны глубокими лиманами, в том числе Тазовского, проникающего на 600 м. в глубь материка, а Енисея на 300 м. Таймырский, Фаддейский и Хатангский заливы представляют собой широкие и глубокие котловины, почти круглый год скованные льдом. Полуостров Таймыр, между Енисеем и Хатангой, представляет собой каменистую тундру.
Правительство осушено Енисеем и его притоками. На 55° с.ш. эта река подходит к Чулыму, притоку Оби, от которого ее отделяет перешеек всего 6 м.по ширине. Часто обсуждалась возможность соединения в этом месте двух великих речных систем Сибири; сложность в том, что Чулымская долина на 440 футов выше другой.
Енисейск богат всевозможными металлами и полезными ископаемыми. Золотой песок появляется в Северо-Енисейской тайге, в районе Кузнецкого Алатау и его отрогов, с бассейнами рек Тубы, Сисима и Черного и Белого Юса, в верховьях притоков Кана и Агула. . Серебряная руда находится в бассейне Абакана, но рудники заброшены. Железная руда встречается почти повсеместно в С. Енисейске, но на Абакане есть только один железорудный завод. Соленые озера распространены.
Железная руда встречается почти повсеместно в С. Енисейске, но на Абакане есть только один железорудный завод. Соленые озера распространены.
Климат, хотя и очень суровый, предлагает большое разнообразие. Минусинские степи отличаются сухим и относительно мягким климатом. В Красноярске (55° 1′ с. ш.) климат более суров, а ветры неприятны. Годовое количество снега настолько мало, что ветры сдувают его в окрестностях города. В городе Енисейске (58° 27 с. ш.) средняя температура ниже точки замерзания, а в Туруханске самый холодный месяц (февраль) в среднем — 24° F.На Таймыре средняя летняя температура едва достигает 45°.
Нагорья Саяна и Алатау густо покрыты лесами из кедра, смоляной сосны, лиственницы, бузины и березы, с рододендронами, Berberis и Ribes; пихта обыкновенная встречается только в нижних и более сухих частях долин. Вершины и склоны гор усыпаны щебнем и валунами, густо покрыты лишайниками и мхами; но есть участки лугов, украшенных цветами, большинство из которых известны в Европе. Однако флора, как правило, бедна, и д-ру Мартьянову после нескольких лет работы удалось собрать только 104 вида фанерогам. 1 С другой стороны, Минусинские равнины и степи Абакана пестрят цветами, разбросанными среди обыкновенных злаков, и в июне и июле Polygala, Dianthus, Medicago, Lathyrus, желтой душистой лилии, и множество других цветов, в основном знакомых в Европе, но достигающих в Енисейске более крупных размеров и большей яркости окраски.Богатый ковер из травы и цветов увенчан высокими белыми цветами Archangelica и Spiraea Ulmaria, и голубыми массами Veronica longifolia. Луга более влажных местностей, окруженные зарослями ивы, тополя, черешни и боярышника, еще более привлекательны из-за их обилия анемонов, фиалок, горечавок и т. д., а также многочисленных лиан, украшающих деревья и кустарники. Всего в списках д-ра Мартьянова насчитывается 760 цветковых и 760 скрытогамных растений.Из низших грибов и паразитических миксомицетов отмечено 1300 видов, а из 823 видов, описанных до сих пор специалистами, не менее 124 оказались новыми.
Однако флора, как правило, бедна, и д-ру Мартьянову после нескольких лет работы удалось собрать только 104 вида фанерогам. 1 С другой стороны, Минусинские равнины и степи Абакана пестрят цветами, разбросанными среди обыкновенных злаков, и в июне и июле Polygala, Dianthus, Medicago, Lathyrus, желтой душистой лилии, и множество других цветов, в основном знакомых в Европе, но достигающих в Енисейске более крупных размеров и большей яркости окраски.Богатый ковер из травы и цветов увенчан высокими белыми цветами Archangelica и Spiraea Ulmaria, и голубыми массами Veronica longifolia. Луга более влажных местностей, окруженные зарослями ивы, тополя, черешни и боярышника, еще более привлекательны из-за их обилия анемонов, фиалок, горечавок и т. д., а также многочисленных лиан, украшающих деревья и кустарники. Всего в списках д-ра Мартьянова насчитывается 760 цветковых и 760 скрытогамных растений.Из низших грибов и паразитических миксомицетов отмечено 1300 видов, а из 823 видов, описанных до сих пор специалистами, не менее 124 оказались новыми. Дальше С. флора сходна по характеру с флорой Сибирской низменности (см. Сибирь). На Таймырском полуострове представлено всего 124 видами цветковых растений.
Дальше С. флора сходна по характеру с флорой Сибирской низменности (см. Сибирь). На Таймырском полуострове представлено всего 124 видами цветковых растений.
Степи Верхнего Енисея были заселены с глубокой древности, и бесчисленные курганов, или курганов, могил, наскальных надписей и плавильных печей сменяющих друг друга жителей разбросаны по всем прериям Абакана и Минусинска.2 Нынешнее население демонстрирует следы всех своих предшественников. В степях и Саянах встречаются многочисленные остатки тюркских и самодийских племен; но некоторые из них значительно сократились в численности. Расчетное население в 1906 году составляло 657 900 человек. Она почти сплошь русская, остальные (около 10%) состоят из самодийцев, татар, тунгусов, якутов, монголов и остяков. Правительство разделено на пять районов, главными городами которых являются Красноярск, Ачинск, Канск, Минусинск и Енисейск.
1 Мартьянов Н., «Материалы для флоры Минусинской области», в Труды Казанского общества естествоиспытателей (xi. 3, 1882).
3, 1882).
2 См. Радлов В., Aus Sibirien (2 тт., Лейпциг, 1880 г.), и Савенков Н., в Известиях Восточно-Сибирского географического общества (XVII, 1887).
Около 1 117 000 акров (0,2%) заняты посевами, в основном под посевами ржи, пшеницы, овса, ячменя и картофеля. Широко развито животноводство, в том числе оленеводство.Большое значение имеет рыболовство, особенно на низовьях Енисея. Соболей теперь не найти, и охотники добывают главным образом белок, лисиц, песцов и медведей. В середине 19 века от 350 000 до 525 000 унций. золота ежегодно добывали в Северном и Южном Енисейске, но к концу века добыча упала до менее 100 000 унций. Соль добывается так же, как и английская соль. Уголь найден на Нижней Тунгуске, у устья Енисея и во многих местах С.правительства. Серебро, медь, свинец, бурый уголь или бурый уголь, каменная соль, графит и слюда существуют в больших количествах, но добываются нерегулярно. Есть несколько ликероводочных заводов. Торговля ведется мехами (экспорт), бакалейными товарами и промышленными товарами (импорт). Прииски Енисейской тайги снабжаются зерном и скотом по реке из Минусинска. области, а солью, спиртом и железом у Ангары. Правительство проходит от З. до В. Сибирской железной дорогой, и приложены значительные усилия для установления регулярного пароходного сообщения между устьем Енисея и З.Европа. За несколько лет до конца XIX века пароходы (например, пароход английского капитана Виггинса) подходили к устью Енисея, ввозя продовольствие и оборудование для золотых приисков. Усилия. предприняты меры по расчистке порогов Ангары, чтобы привести Байкал в пароходное сообщение с Енисеем. Однако из-за мелководья мелких притоков Енисея канал, соединяющий Енисей с Обью, оказался не таким пригодным, как ожидалось. (П.А. К.; Дж. Т. Б.Э.)
Прииски Енисейской тайги снабжаются зерном и скотом по реке из Минусинска. области, а солью, спиртом и железом у Ангары. Правительство проходит от З. до В. Сибирской железной дорогой, и приложены значительные усилия для установления регулярного пароходного сообщения между устьем Енисея и З.Европа. За несколько лет до конца XIX века пароходы (например, пароход английского капитана Виггинса) подходили к устью Енисея, ввозя продовольствие и оборудование для золотых приисков. Усилия. предприняты меры по расчистке порогов Ангары, чтобы привести Байкал в пароходное сообщение с Енисеем. Однако из-за мелководья мелких притоков Енисея канал, соединяющий Енисей с Обью, оказался не таким пригодным, как ожидалось. (П.А. К.; Дж. Т. Б.Э.)
Центральная система бронирования Академсервис — бронирование отелей онлайн
ФИО: Минусинск
Страна: Россия
Регион: Красноярский край
Часовой пояс: UTC+7
Площадь: 17,7 кв. км
км
Население: 0,068 млн (2014)
Год основания: 1739
Климат: континентальный
Средняя температура: от -26°C до -10°C зимой, от 9°C до 27°C летом
Код города: +7 (39132)
Аэропорты: Абакан
Вокзалы: Минусинск
Автовокзалы: Минусинск
Культовые сооружения: Православные: Спасо-Преображенский собор, Св.часовня Пантелеймона; Протестантская: Церковь евангельских христиан-баптистов
Архитектурные памятники: Дом купца Вильнера, Дом купца Белова, Гостиный двор, Казначейские склады, Государственные учреждения, Здание Госсклада Винной Монополии, Здание Драматического Театра
Статуи и памятники: А.Д. Кравченко, П.Е. Щетинкин, С.К. Сургуладзе, С.И. Кретов, Н.М. Мартьянов, Ветераны ВОВ, Жертвы политических репрессий, Памятный знак «Жертвам Чернобыльской и других радиационных катастроф»
Музеи: N. Областной краеведческий музей им. М. Мартьянова, Галерея изобразительных искусств, Музей «Автомобили и автомашины СССР», Музей декабристов
Областной краеведческий музей им. М. Мартьянова, Галерея изобразительных искусств, Музей «Автомобили и автомашины СССР», Музей декабристов
Дома-музеи: Г.М. Кржижановский и В.В. Мемориальный музей-квартира Старкова
Театры: Драматический театр
Природные объекты и парки: Городской парк культуры и отдыха
Спортивные объекты и клубы: Стадион «Электрон», Стадион «Строитель», Гимназия «Строитель», Ю.Спорткомплекс имени В. Шумилова
Площадки для выступлений: Городской Дом культуры
Развлекательные объекты: Кинотеатр «Альянс II», Детский развлекательный центр «Егоза»
Рестораны национальной кухни: Блин Ком (русская), Айвенго (европейская), Pizza mia (итальянская), Оки-Доки (японская)
Томат «Минусинские»
УПАКОВКА: Пакет на молнии 12–15 СЕМЯН
Крупные розовые помидоры «бычье сердце». Устойчив к плохим погодным условиям.
Устойчив к плохим погодным условиям.1. Оригинальное название: Неизвестно. Это старый сорт, происходящий из Минусинска и названный в честь этого города.
2. История: Минусинские сорта известны в Красноярском крае с конца 19 века. Впервые их ввели приговоренные к ссылке декабристы. Район (Минусинская котловина) был хорош для выращивания овощей. В 1913 году местному селекционеру томатов Старухину удалось вырастить плод весом 26 унций.(745 г). Этот сорт получил название Минусинские или Минусинские.
3. Характеристики растений: Раннеспелый. В теплице более успешно. Индетерминантный куст среднего размера, высотой 130-160 см. Тонкие стебли, обычный лист. Кисть регулярная, по 3-4 крупных плода на кисти.
4. Инструкции по дрессировке : Желательно прививать куст к двум или трем основным стеблям. Растение выглядит слабым, поэтому необходимо подвязывать и обрезать.
5. Форма и вес: Большие и круглые плоды сердцевидной формы с плоскими заостренными концами. Иногда у конца стебля имеется несколько неглубоких ребер. Большой вес. Плоды могут весить до 10,5-14 унций. (300-400 г) и даже больше. Иногда они могут весить до 35 унций. (1 кг) при контролируемом цветении и использовании хороших удобрений.
Иногда у конца стебля имеется несколько неглубоких ребер. Большой вес. Плоды могут весить до 10,5-14 унций. (300-400 г) и даже больше. Иногда они могут весить до 35 унций. (1 кг) при контролируемом цветении и использовании хороших удобрений.
6. Цвет: Ярко-розовый, очень красивый. В разрезе тоже розовый.
7. Профиль вкуса: Чудесный вкусный сладкий вкус розовых помидоров. Такими помидорами невозможно наесться. Мякоть глянцевая и мясистая, но при этом очень сочная.
8. Кулинарное применение: Идеально подходит для салатов – много салатов! Идеально подходит для консервирования превосходных соков. Плохо хранится после сбора урожая. Плоды имеют тенденцию трескаться, когда растения получают слишком много воды слишком быстро.
Томаты Минусинские — устойчивый сорт с крупными и вкусными плодами. Прежде всего, они устойчивы к холоду!
Туристическая фирма «Сибирь»
Территория в основном расположена в Восточной Сибири, в бассейне реки Енисей.
 На севере район омывается Карским морем и морем Лаптевых. Географический центр России расположен в Красноярском крае.
На севере район омывается Карским морем и морем Лаптевых. Географический центр России расположен в Красноярском крае. Площадь области составляет 2339,7 кв. км (13,6% территории РФ). Население 3095,1 тыс. человек. Столица и крупнейший город — Красноярск. Время: московское плюс 4 часа.
Область охватывает 3 климатические зоны: арктическую, субарктическую и умеренную. Центральная часть области, в основном равнинная, характеризуется жарким летом и продолжительной холодной зимой и быстрой сменой температур.Для юга области характерно теплое лето и умеренная малоснежная зима. Средняя температура января -36С на севере, -17С на юге, июля — +10С и +20С соответственно.
Населенные пункты Красноярского края
г. Красноярск
Расположен на границе Восточных Саян, отрогов Енисейского кряжа и Западно-Сибирской равнины, по обоим берегам реки Енисей. В 1628 году казак Андрей Дубенский основал имение Красный Яр.Первыми жителями усадьбы были казаки, и до 19 века город был центром казачьей общины.
В 1822 г. Красноярск стал центром Енисейской губернии. В конце 19 века через Красноярск прошла Транссибирская магистраль, и город начал бурно развиваться.
В 1934 году Красноярск стал центром Красноярского края. В первые годы Великой Отечественной войны в Красноярск был эвакуирован ряд крупных заводов, что предопределило путь развития города как промышленного центра.Современный Красноярск вытянут с запада на восток на 18-20 км по левому берегу и на 25 км по правому берегу Енисея.
Площадь города составляет 348 кв. км. Население — 877 тыс. жителей. Красноярск сегодня — административный, экономический и культурный центр, крупнейший город Восточной Сибири, очень гостеприимный для приезжих.
г. Минусинск
Город расположен в центральной части Минусинской котловины, в 430 км от Красноярска. Население 74 человека.3 тысячи. Город входит в список исторических мест России. Он очень интересен своей архитектурой. В старой части города сохранились деревянные дома, построенные на каменном фундаменте, бывшие купеческие дома, окна, украшенные деревянными изделиями, лавки с окованными железом дверями и металлическими решетками.
г.Дивногорск
Расположен в 40 км от Красноярска на правом берегу Енисея. Это город работников Красноярской ГЭС.Название городу дали дивные Саянские горы, раскинувшиеся на противоположном, левом берегу Енисея. Дивногорск – одно из самых живописных и экологически чистых мест области. Население составляет 30,3 тыс. человек.
г. Енисейск
Расположен на берегу реки Енисей в 330 км к северу от Красноярска. Население составляет 21,3 тыс. жителей. Енисейск называют «музеем под открытым небом», в него входят 94 здания, являющиеся памятниками архитектуры прошлого.Комплекс Спасского монастыря и 4 церквей являются уникальными образцами сибирского барокко, построенными в середине и во второй половине 18 века. Город отличается рядом зданий, построенных в классическом стиле, модерне и эклектике.
село Овсянка
Расположен в 26 км от Красноярска на берегу реки Енисей. Население 4,3 тыс. человек. Выдающийся русский писатель Виктор Астафьев родился и жил в Овсянке (1994-2001). Главные достопримечательности поселка – дом-музей Астафьева, библиотека-музей и собор Св.Инокентская Иркутская церковь.
Главные достопримечательности поселка – дом-музей Астафьева, библиотека-музей и собор Св.Инокентская Иркутская церковь.
село Сушенское
Поселок расположен на юге Красноярского края, в 488 км от Красноярска. Население — 22 тыс. чел. Село основано в 1744 году. В 19 веке было местом политической ссылки. В 1870-1900 годах сюда был сослан Владимир Ленин.
Главной достопримечательностью поселка является государственный историко-этнографический музей-заповедник «Шушенское», в котором представлен быт сибирского села 19-20 веков.
д. Курагино
Центр крупнейшего района на юге Красноярского края, он имеет уникальное географическое положение: степные, частично лесостепные, высокогорные районы.Деревня расположена на берегу реки Тубы, протекающей через весь район.
Памятников архитектуры 201, из них 21 охраняется государством.
Деревня Курагино была основана в 1626 году. В 17 веке здесь была тубинская татарская деревня. Русские, после захвата Тубинского княжества, назвали это место Курагинском, по имени князя Кураги.
В 1834 г. была образована Курагинская губерния, в апреле 1924 г. образована Курагинская область. Городской поселок с 1961 года.Население составляет 15 тысяч жителей.
Экскурсионный
Красноярский краевой музей (Красноярск)
Он был открыт в 1889 году. Для значительного увеличения коллекции в 1913-1914 годах было построено новое здание по проекту известного художника и архитектора Л.А. Чернышева. Он был открыт только в 1930 году. В 2001 году, после 14-летней реконструкции, обновленный и расширенный музей вновь открылся для горожан и гостей города. Сейчас Красноярский краеведческий музей — один из лучших в России.Основные коллекции – археологические, этнографические, естественнонаучные, нумизматические, художественные, вещевые, фото- и документальные материалы. Фонды музея состоят из 400 тысяч единиц хранения.
В.И. Дом-музей Сурикова, выдающегося русского художника-передвижника (1848-1916).
В своих масштабных полотнах он посвящен переломным моментам русской истории («Утро стрельцов», «Меньшиков в Березове», «Боярыня Морозова» и др. ). главными героями были простые люди.Музей был открыт в 1948 году в доме художника, где он родился и жил до отъезда на учебу в Петербург. Дом был построен в 1830 году отцом художника. Сохранился внешний вид дома и внутреннее убранство десяти комнат, а также конюшня, русская баня и сарай. В коллекции музея-усадьбы около 100 оригинальных произведений искусства, в том числе малоизвестные акварели, а также личные вещи художника.
). главными героями были простые люди.Музей был открыт в 1948 году в доме художника, где он родился и жил до отъезда на учебу в Петербург. Дом был построен в 1830 году отцом художника. Сохранился внешний вид дома и внутреннее убранство десяти комнат, а также конюшня, русская баня и сарай. В коллекции музея-усадьбы около 100 оригинальных произведений искусства, в том числе малоизвестные акварели, а также личные вещи художника.
Заповедник «Столбы» (г. Красноярск)
Находится в 3 км от Красноярска.Заповедник основан в 1925 году. Площадь 47,2 га, наиболее посещаемая часть заповедника 1400 га. Это паломничество скалолазов и альпинистов. В гористой местности, среди светлого соснового леса из лиственниц и кедров, миллионы лет дожди и зимы, мороз и солнце вырезали из камней причудливые фигуры. Контуры фигур сильно напоминают птиц, животных и людей.
Красноярская ГЭС
Строился с 1959 по 1971 год.Красноярская ГЭС – вторая по мощности в России. Плотина высотой 124 м образует большой водоем протяженностью 380 км и площадью 2 тыс. кв. км. Гидроагрегаты состоят из бетонной плотины, цеха ГЭС и уникального наклонного элеватора. В мастерской находится музей гидроэлектростанции.
кв. км. Гидроагрегаты состоят из бетонной плотины, цеха ГЭС и уникального наклонного элеватора. В мастерской находится музей гидроэлектростанции.
Государственный историко-этнографический музей-заповедник «Шушенское» (с. Шушенское)
Уникальный музейный комплекс под открытым небом.Музей основан в 1937 году. В начале 1970-х годов он был ориентирован на отображение быта, традиций и обычаев, аттракционов, этнографии сибирской деревни XIX-XX веков. А до 1993 года это был музей-заповедник «Сибирская ссылка Ленина»
.
Музей занимает 7 га с экспонатами под открытым небом и 29 усадьбами, каждая из которых является музеем различных крестьянских ремесел и промыслов. Очень интересно увидеть магазин с разными вещами в нем, деревенскую кузницу, трактир и дома, связанные с пребыванием здесь Ленина, с которых и началась экспозиция музея.
Минусинский краеведческий музей им. Н. Мартьянова (Минусинск)
Музей создан в 1877 году химиком и коллекционером Николаем Мартьяновым, который собрал уникальные археологические находки, обнаруженные в Минусинской котловине, имеющие мировое значение. В 1900 г. на Всемирной выставке в Париже музей был удостоен серебряной медали.
В 1900 г. на Всемирной выставке в Париже музей был удостоен серебряной медали.
Сотрудниками музея собран богатейший фонд, насчитывающий более 200 тысяч экспонатов; библиотека насчитывает более 100 тысяч книг.В музее собраны зеркала китайского средневековья, китайские монеты 5-19 веков, произведения искусства Тувы и Хакасии.
Что к чему
Ергаки — горный массив
Чудесное место Восточной Сибири. Он расположен между городом Абакан и городом Кызыл. «Еграки» в переводе с древнетюркского означает «пальцы». На самом деле там много скал, напоминающих растопыренные пальцы и называемых Зубом Дракона, Каменным городом, Спящим Саян, Висячим камнем и т.д.Регион известен как культовое место для туристов.
Железнодорожная дорога «Мужество» Абакан-Тайшет.
Это кратчайшая железнодорожная магистраль, соединяющая важнейшие промышленные центры Западной и Восточной Сибири.
Железная дорога проходит по сложнейшим горным хребтам Саянских гор. 647 км пути имеют 49 станций, 9 тоннелей протяженностью более 10 км, 5 огромных виадуков, два из которых не имеют себе равных в мире, 78 мостов. Строительство началось в 1958 году, а 9 декабря 1965 года был подписан Акт приемки железной дороги, введенной в эксплуатацию.
647 км пути имеют 49 станций, 9 тоннелей протяженностью более 10 км, 5 огромных виадуков, два из которых не имеют себе равных в мире, 78 мостов. Строительство началось в 1958 году, а 9 декабря 1965 года был подписан Акт приемки железной дороги, введенной в эксплуатацию.
Минусинская котловина
Минусинская котловина — крупный район, расположенный на юге Красноярского края и в Хакасии. Он известен своей плодородной почвой. Здесь также много археологических памятников, курганов, стоянок древних городов и поселений, остатков древних плавильных печей.
Партизанское движение в годы Гражданской войны (1917-1920 гг.)
В 1918-1920 гг. на юге Красноярского края развернулось движение рабочих и крестьян против интернациональной военной интервенции и войск Колчака.Самый ранний и самый мощный центр был создан в Степно-Баджейской губернии.
Руководителем восстания был Кравченко А. — агроном по профессии и военный по наклонностям. На территории, контролируемой партизанами, была основана Степно-Баджейская республика.
Зимой 1918-1919 гг. колчаковские власти предприняли карательные акции по ликвидации партизанского движения. Туда были направлены отряды милиции Колчака, казачьи части, итальянские, сербские, чехословацкие легионеры.Во второй половине 1919 года численность сил, брошенных на подавление степно-баджейского партизана, достигла 12 тысяч человек. Каратели расправились с местным населением в партизанском районе с беспричинной жестокостью. Во всей Степно-Баджейской губернии не осталось ни одного дома. Партизанская армия была вынуждена отступить в район Минусинска и уйти на юг к Белоцарску.
Фитобочка
«Кедровая фитобочка» (фитобочка) — вид бани. Издревле на Руси известно благотворное влияние паров бани на организм.Основная особенность, используемая в «кедровой фитобочке», заключается в том, что голова пациента находится вне ствола, а пар, насыщенный различными природными концентрированными веществами, поступает в ствол под давлением. При высоких температурах начинается сильное потоотделение, расширяются кровеносные сосуды, улучшается кровоснабжение кожи, мышц и суставов. Процедура занимает 5-15 минут.
Процедура занимает 5-15 минут.
Хребет Оглахты — Центр всемирного наследия ЮНЕСКО
Описание
Хребет Оглахты расположен на левобережье Енисея (Красноярское водохранилище) и представляет собой систему низменных хребтов с большим количеством выходов пород девонского красного песчаника, обеспечивающих, с одной стороны, живописный ландшафт, а во всем его использование человеком. тысячи лет, с другой стороны.Тысячи наскальных рисунков, относящихся к разным историческим периодам, на вертикальных выходах горных пород и обломках скал свидетельствуют об изменении среды обитания, типов земледелия, материальной и духовной культуры народов, населявших эту территорию. Из этого же камня возведены внушительные погребальные сооружения, встречающиеся в районах предгорий и межгорных степных долин, и огромное укрепление, окаймляющее горы с южной стороны. Номинируемая территория представляет большой научный интерес с точки зрения природного наследия: разнообразием ландшафта; редкие эндемичные растения; 5 видов рептилий, 148 видов птиц и 44 вида млекопитающих. Частично территория входит в состав участка «Оглахты» государственного природного заповедника «Хакасский», зарегистрированного в 1991 году. Ключевой задачей заповедника является сохранение и защита степных биоценозов, местообитаний редких эндемичных растений и редких видов животных, скальных птиц ансамбли, памятники историко-культурного наследия.
Частично территория входит в состав участка «Оглахты» государственного природного заповедника «Хакасский», зарегистрированного в 1991 году. Ключевой задачей заповедника является сохранение и защита степных биоценозов, местообитаний редких эндемичных растений и редких видов животных, скальных птиц ансамбли, памятники историко-культурного наследия.
Природное наследие
Геология и рельеф
Низкогорный ярко выраженный моноклинальный Оглахтинский хребет, расположенный на левом берегу Енисея (Красноярское водохранилище) ниже устья реки Тубы (правый приток Енисея), разделяет Минусинскую (Южно-Минусинскую) и Сыда-Ербинскую (Центрально-Минусинскую) реки. Minusinsk) котловины обширной Минусинской котловины в горах Южной Сибири.Формирование депрессии происходило в девонский (410—360 млн лет назад) период. Это было время устойчивого многолетнего поверхностного тектонического оседания. И в то же время складчатый скальный фундамент был расколот на отдельные блоки, в результате чего разделение некоторых частей сохранилось до сих пор. Древние породы, образующие Минусинскую котловину, неоднократно подвергались тектоническим воздействиям и образовывали как складчатые, так и глыбовые горы. Под воздействием внешних факторов (таких как ветер, вода, колебания температуры) горы последовательно размывались, при этом более плотные породы размывались значительно медленнее.Так было создано то разнообразие рельефа, которое характерно для этого края: обширные равнины, холмы, хребты и невысокие горы. Ныне невысокие горы, сложенные девонским красно-бурым глинистым камнем, возвышаются над окружающей местностью, что является характерной чертой хакасских ландшафтов.
Древние породы, образующие Минусинскую котловину, неоднократно подвергались тектоническим воздействиям и образовывали как складчатые, так и глыбовые горы. Под воздействием внешних факторов (таких как ветер, вода, колебания температуры) горы последовательно размывались, при этом более плотные породы размывались значительно медленнее.Так было создано то разнообразие рельефа, которое характерно для этого края: обширные равнины, холмы, хребты и невысокие горы. Ныне невысокие горы, сложенные девонским красно-бурым глинистым камнем, возвышаются над окружающей местностью, что является характерной чертой хакасских ландшафтов.
Хребет Оглахты сложен осадочными породами нижнего карбона и верхнего девона: песчаниками, алевролитами, туффитами, известняками, доломитами. Его холмистый рельеф перемежается куэстовыми хребтами с крупными выходами скал и небольшими долинами.Эта территория представляет собой прекрасный пример куэстового рельефа (образованного за счет наклонного залегания разноплотных пород). Куэсты, вытянутые в широтном направлении, имеют крупные пологие складки, осложненные флексурами и флексурообразными изгибами слоев. Они образуют систему длинных параллельных возвышенностей, между которыми развиваются куэстовые овраги. Эоловые процессы играют ключевую роль в экзогенных рельефообразующих процессах, благодаря им получили широкое распространение не только куэсты, но и котловины дефляции и потерянные горы.На всей территории отчетливо наблюдаются избирательные процессы выветривания и денудации, а также жесткая зависимость морфологии поверхности от геологического строения. Относительные высоты 100-150 м. Высшая точка (580 м над уровнем моря) — гора Оглахты. Ареал простирается на 15 км с юга на север.
Куэсты, вытянутые в широтном направлении, имеют крупные пологие складки, осложненные флексурами и флексурообразными изгибами слоев. Они образуют систему длинных параллельных возвышенностей, между которыми развиваются куэстовые овраги. Эоловые процессы играют ключевую роль в экзогенных рельефообразующих процессах, благодаря им получили широкое распространение не только куэсты, но и котловины дефляции и потерянные горы.На всей территории отчетливо наблюдаются избирательные процессы выветривания и денудации, а также жесткая зависимость морфологии поверхности от геологического строения. Относительные высоты 100-150 м. Высшая точка (580 м над уровнем моря) — гора Оглахты. Ареал простирается на 15 км с юга на север.
Сильно фрагментированный рельеф, переменная крутизна склонов и экспозиция обеспечивают большое разнообразие ландшафтов на этой сравнительно небольшой территории.
Восточная граница ареала связана с левобережной полосой Енисея (с 1970 г. Красноярское водохранилище). Здесь можно увидеть высокие живописные скалы, круто обрывающиеся в реку. Протянувшиеся на северо-запад в пределах хребта глубокие овраги в нескольких местах прорезают фронт обрыва. Один из них представляет собой довольно широкую долину, разделяющую ареал на четко выраженные северную и южную части.
Здесь можно увидеть высокие живописные скалы, круто обрывающиеся в реку. Протянувшиеся на северо-запад в пределах хребта глубокие овраги в нескольких местах прорезают фронт обрыва. Один из них представляет собой довольно широкую долину, разделяющую ареал на четко выраженные северную и южную части.
Климат. Географическое положение, орографические особенности и атмосферная циркуляция оказывают существенное влияние на формирование климата Оглахтинского хребта. Климат этой территории, как и климат горных степей Минусинской котловины в целом, континентальный, с холодной продолжительной зимой (пять месяцев) и коротким жарким летом.Для него характерны не только месячные, но и суточные резкие колебания температуры. Годовое количество осадков колеблется от 250 до 300 мм. Средняя температура января в пределах –16–20°С, июля в пределах +18–20°С. Минимальная температура достигает –40–45°С, а максимальная до +35–40°С. Минусинская котловина превосходит по количеству солнечных дней в году Крым.
Почвы. Почвенный покров характеризуется преобладанием среднегумусных, среднемощных южных и обыкновенных черноземов, покрывающих пологие склоны.Черноземы малогумусные, как маломощные, так и среднемощные, являются постоянным компонентом почвенно-покровного комплекса. Малогумусные черноземы в юго-западной части района развиты на красноцветных и преимущественно щебнистых породах. В приенисейской холмистой части степей распространены щебнистые почвы. Все почвы тяжелые. Почвы северных лесистых склонов дерново-подзолистые. Некоторые почвы Оглахтинского хребта занесены в Красную книгу почв и Кадастр особо ценных почвенных объектов России, например чернозем обыкновенный среднемощный среднесуглинистый на лессовидных суглинках (не обрабатывался 30 лет).Почвы этой территории представляют собой специфические природно-исторические образования, которые могут служить эталонами для сравнения с измененными в результате деятельности человека, а также для оценки ущерба почвам и экосистемам с последующей разработкой методов восстановления.
Флора. В хребте Оглахты и прилегающих к нему степных территориях произрастают малоплодородные и эндемичные растения, такие как лабазник минусинский ( Hedysarum minussinense ), мартьянов теплолистный ( Artemisia martjanovii ), остролистник ( Oxytropis encludens ) и другие.Уже зарегистрировано 234 вида высших видов сосудистых растений. Большую часть территории занимают низкокустарничковые и высококустарниковые степи и их петрофитные скопления. Реже встречаются луговые степи и остепненные сухолуга. Северные склоны большей частью покрыты березовыми лесами и степными кустарниками.
Настоящие мелкокустарниковые степи встречаются как на равнинных, так и на склоновых участках на каштановых и южно-черноземных почвах. Типичные виды растений: 1) Кусты: Caragana pygmaea. 2) Засухоустойчивые злаки образуют травяной покров: Festuca pseudovina, Koeleria cristata, Poa sibirica, Cleistogenes squarrosa, Carex duriuscula, Carex pediformis. Из разнотравья обычные Veronica incana, Aster alpinus, Heteropappus altaicus subsp. altaicus, Artemisia frigida, Artemisia glauca, Leontopodium ochroleucum subsp. campestre, Hedysarum gmelini и довольно большое количество растений семейств Oxytropis и Astragalus.
Из разнотравья обычные Veronica incana, Aster alpinus, Heteropappus altaicus subsp. altaicus, Artemisia frigida, Artemisia glauca, Leontopodium ochroleucum subsp. campestre, Hedysarum gmelini и довольно большое количество растений семейств Oxytropis и Astragalus.
Истинные высоко- Кустарниково- Кустарниковые степи встречаются на остепненных участках склонов западной и восточной экспозиции и переходят на южные склоны в лесостепных районах. Типичными видами растений являются: 1) Травяной покров: преобладают Stipa capillata или Helictotrichon Desertorum . Другие виды растений следующие: POA StepPapa , , Agropyron Cristatum , Флиоида Флюи , Carex Pediformis , Dianthus Versicolor , Galium Verum Delphinium Grandiflorum , , Artemisia Glauca , Medicago Falcata , Onobrychis Arenaria , Aster Alpinus, гетеропаппус Altaicus Subsp. алтайский.
алтайский.
Полупустынные степи не имеют широкого распространения и встречаются фрагментарно на южных склонах. Растительность редкая и низкая. Покрытие растительностью не превышает 40%. Типичные виды растений: 1) Кусты: Caragana pygmaea . 2) Травяной покров: Panzerina lanata subsp. argyracea , Thymus Minussinensis , Artemisia Frigida , Ceratoides Papposa , Kochia Laniflora и низкая трава.
Луговые степи и степные сухие луга находятся на северных склонах. Типичные виды растений: 1) Trash Cover: в основном Iris Ruthenica , Флеоида , POA StepPapa , Bromopsis Inermis , менее часто STIPA Pennata . Травы в том числе: Bupleurum multinerve , Дельфиниум grandiflorum , земляника зелёная , кровохлёбка лекарственная , люцерна серповидная , Astragalus danicus , Dracocephalum ruyschiana , , Phlomis Tuberosa , Scabiosa Ohroleuca , Ligularia Glauc и Растения Potentilla, Geranium , Семьи Thalictrum и другие .
Каменистые степи находятся на крутых склонах южных экспозиций. Типичными видами растений являются: 1) Древесные породы: лиственничные редколесья часто встречаются на южных склонах ( Larix sibirica ). 2) Травяной покров: кроме обычных степных видов имеются реликтовые: Patrinia sibirica , Kobresia filifolia , Gypsophila patrinii , 9031us. , хамеродосово ereca, Orostachys зртоза , Arctogeron gramineum , Goniolimon speciosum , Androsace максимумы , Хвойник Односемянный , Спаржа pallasii , Sedum aizoon, Sedum hybridum , Phlox sibirica , Onosma simplicissima .
Древесно-кустарниковая растительность встречается на северных склонах, в мелколесьях и на опушках, и лишь отдельные участки покрыты мелколесьем. Типичными видами растений являются: 1) древесные виды: Betula pendula и Larix sibirica, реже Populus nigra . 2) кусты: в основном Spiraea Hypericifolia, Spiraea Media, Spiraea Chamaedryforfylia , Cotoneaster Melanocarpus , Rosa acibrici, Garagana arborescens , реже Padus Avium , Crataegus Sanguinea , , , , , , Grossularia acicularis , Lonicera tatarica, Atraphaxis frutescens.3) Грасс Крышка: Bupleurum multinerve , Дельфиниум grandiflorum , земляника зелёная , кровохлёбка лекарственная , люцерна серповидная , астрагал danicus , Dracocephalum ruyschiana , зопник Tuberosa , Scabiosa ohroleuca , Ligularia GLAUCA и растения otentilla, герань , Thalictrum семей и другие ( Hemerocallis Lilio-Asphodelus , Trollius asiaticus , Anemone sylvestris и другие .
Типичными видами растений являются: 1) древесные виды: Betula pendula и Larix sibirica, реже Populus nigra . 2) кусты: в основном Spiraea Hypericifolia, Spiraea Media, Spiraea Chamaedryforfylia , Cotoneaster Melanocarpus , Rosa acibrici, Garagana arborescens , реже Padus Avium , Crataegus Sanguinea , , , , , , Grossularia acicularis , Lonicera tatarica, Atraphaxis frutescens.3) Грасс Крышка: Bupleurum multinerve , Дельфиниум grandiflorum , земляника зелёная , кровохлёбка лекарственная , люцерна серповидная , астрагал danicus , Dracocephalum ruyschiana , зопник Tuberosa , Scabiosa ohroleuca , Ligularia GLAUCA и растения otentilla, герань , Thalictrum семей и другие ( Hemerocallis Lilio-Asphodelus , Trollius asiaticus , Anemone sylvestris и другие . )
)
Животный мир типичный степной и представлен дикой природой открытых пространств, древесно-кустарниковой растительностью и выходами скал. Зарегистрировано 44 вида млекопитающих, 148 видов птиц, 5 видов рептилий и 26 видов рыб (в Красноярском водохранилище).
Характерные виды животных: 1) A земноводные: остромордая лягушка ( Rana arvalis). 2) Рептилии: прыткая ящерица ( Lacerta agilis), среднеазиатская гадюка (Gloydius halys), обыкновенная ящерица (Zootoca vivipara), обыкновенная северная гадюка (Vipera berus). 3. caudatus), белая синица (Parus montanus), чернозобый дрозд (Turdus atrogularis), черноголовый чекан (Saxicola torquata), желтобровая камышевка (Phylloscopus inornatus), алый грос обыкновенный перепел ( Coturnix coturnix ), куропатка ( Perdix daurica ), тетерев ( Lyrurus tetrix ) . 4) Mammals: млекопитающих: Common Shrew Sorex ( Sorex Araneus), SICTISTA (Sicista Subtilis), DWARF Hamster (Phodopus Sungorus), Steppe Lemming (Lagurus Lagurus), полевая мышь (Microtus arvalis), водяная полевка (Arvicola terrestris), узкочерепная полевка (Microtus gregalis), полевая мышь (Apodemus agrarius), восточноазиатская мышь (Apodemus peninsulae), домовая мышь (Mus musculus ), длиннохвостый суслик (Spermophilus undulates), барсук (Meles amurensis), заяц-русак (Lepus europaeus), лисица (Vulpes vulpes), волк (Canis lupus), сибирская косуля (Capreolus pygargus).
4) Mammals: млекопитающих: Common Shrew Sorex ( Sorex Araneus), SICTISTA (Sicista Subtilis), DWARF Hamster (Phodopus Sungorus), Steppe Lemming (Lagurus Lagurus), полевая мышь (Microtus arvalis), водяная полевка (Arvicola terrestris), узкочерепная полевка (Microtus gregalis), полевая мышь (Apodemus agrarius), восточноазиатская мышь (Apodemus peninsulae), домовая мышь (Mus musculus ), длиннохвостый суслик (Spermophilus undulates), барсук (Meles amurensis), заяц-русак (Lepus europaeus), лисица (Vulpes vulpes), волк (Canis lupus), сибирская косуля (Capreolus pygargus).
В этом районе обитают такие редкие млекопитающие, занесенные в Красную книгу России, как степной хорек (Mustela eversmanni), редких птиц, занесенных в Красную книгу России: сапсан ( Falco peregrinus ), беркут ( Aquila chrysaetos ), степной орел ( Aquila nipalensis ), балобан ( Falco Cherryg ) и могильник ( Aquila heliaca ).
Культурное наследие
Несомненная ценность номинируемой территории состоит в том, что природный ландшафт богат следами своего культурного развития на протяжении нескольких исторических эпох.
Район Минусинской котловины был освоен еще в каменном веке. Первобытные люди охотились здесь на крупных млекопитающих, некоторые виды которых вымерли, а другие сменили среду обитания. В дальнейшем территория на протяжении тысячелетий заселялась все новыми и новыми народами, одна культура сменялась другой, менялись хозяйственные и культурные типы, способы взаимодействия человека и природной среды. Здесь, на территории Минусинской ложбинной тайги, веками взаимодействовали охотники, рыболовы из Северной Азии и народы из горно-степных зон Средней Азии.Они создали своеобразный культурный синтез, проявившийся в особенностях хозяйственной адаптации разных групп населения (мобильных лесных охотников и рыболовов, оседлых земледельцев и металлургов, кочевых скотоводов), в функционировании внутренних и внешних связей разных сообществ. и способы их сообщения, включая создание эффектных, видимых, идентифицирующих культурные объекты, такие как курганы, стелы, наскальные рисунки в ландшафте.
Оглахтинский хребет и прилегающие к нему степные участки – это лишь небольшая часть Минусинской котловины, но здесь сосредоточены сотни курганов и тысячи наскальных рисунков, а также другие археологические и исторические артефакты (поселения, культовые места, городища, каменоломни).Они имеют длительную историю изучения (с середины XVIII века) и хорошо разработанную хронологию от эпохи палеолита до Нового времени и Нового времени. Наиболее представительными из них являются афанасьевская энеолитическая культура (IV– 1— пол. III тыс. до н. э.), окуневская культура, эпоха ранней бронзы (середина III — начало II тыс. до н. э.), карасукская культура, эпохи поздней бронзы (2 — пол. II — начало I тыс. до н.э.) и культуры раннего железного века: тагарская культура (9-3 вв. до н.э.), тесинская культура (2-3 вв. вв. н.э.) и таштыкской культуры (1–7 вв. н.э.).Впечатляющие каменно-земляные курганы тагарской культуры с оградами и высокими угловыми и промежуточными плитами являются неотъемлемой чертой исторического ландшафта Минусинских степей. Уникальные погребальные маски, мумифицированные останки, сохранившиеся предметы из органических материалов (ткань, мех, дерево), найденные в могильниках таштыкской культуры в Оглахтах, ныне украшают залы Государственного Исторического музея в Москве и Государственного Эрмитажа в Санкт-Петербурге. . Грандиозный памятник I-нач. II тысячелетии нашей эры находится знаменитая Оглахтинская «крепость», руины которой окружают огромную территорию в несколько десятков квадратных километров.Это сооружение состоит из горизонтально уложенных плит песчаника, некоторые остатки стены достигают 6-8 м в ширину и 1 м в высоту, снаружи можно обнаружить следы рва, окружающего стену, в некоторых сохранились «бастионы». места. Стена возводилась многими поколениями, вероятно, сначала для защиты от гуннских, а затем и от уйгурских нашествий.
вв. н.э.) и таштыкской культуры (1–7 вв. н.э.).Впечатляющие каменно-земляные курганы тагарской культуры с оградами и высокими угловыми и промежуточными плитами являются неотъемлемой чертой исторического ландшафта Минусинских степей. Уникальные погребальные маски, мумифицированные останки, сохранившиеся предметы из органических материалов (ткань, мех, дерево), найденные в могильниках таштыкской культуры в Оглахтах, ныне украшают залы Государственного Исторического музея в Москве и Государственного Эрмитажа в Санкт-Петербурге. . Грандиозный памятник I-нач. II тысячелетии нашей эры находится знаменитая Оглахтинская «крепость», руины которой окружают огромную территорию в несколько десятков квадратных километров.Это сооружение состоит из горизонтально уложенных плит песчаника, некоторые остатки стены достигают 6-8 м в ширину и 1 м в высоту, снаружи можно обнаружить следы рва, окружающего стену, в некоторых сохранились «бастионы». места. Стена возводилась многими поколениями, вероятно, сначала для защиты от гуннских, а затем и от уйгурских нашествий.
Наскальное искусство является важнейшим культурным компонентом территории Оглахтинского хребта. Описанные выше особенности рельефа давали древним художникам множество удобных «холстов»: как здесь повсюду можно встретить выходы красного девонского песчаника, так повсюду можно встретить и рисунки на скалах.Изображения выполнены с применением всех известных техник — выбивки, гравировки, шлифовки, росписи. Большая часть росписей расположена на вертикальных плоскостях, но некоторые из них встречаются на каменных блоках, лежащих на склонах, а также на плитах, использовавшихся при строительстве курганов (некоторые изображения относятся к курганному строительству и связаны с погребальными обрядами во время остальные были сделаны следующими поколениями). Наскальные рисунки сгруппированы в разных частях хребта. Особенно богаты такими рисунками скалы по берегу Енисея и юго-западный склон горы «Сорок Зубьев».Каждое из таких скоплений само по себе является очень важным памятником наскального искусства, но все 12 известных в настоящее время живописных скоплений образуют крупнейший комплекс Минусинской котловины — региона, известного как один из самых богатых в мире культурных ареалов наскального искусства.
Историко-культурная ценность этого комплекса не только в количестве источников, но и в их разнообразии, репрезентативности, наличии как типичных, так и уникальных сюжетов, образов и стилей. Оглахтинские петроглифы представляют собой все культурно-хронологические периоды (не менее девяти), выделенные в древнем искусстве Южной Сибири, за исключением уникальных групп, объяснить которые непросто.Ряд изображений выполнен на очень высоком художественном уровне, поэтому они имеют большое эстетическое значение. Наскальное искусство хребта Оглахты является ценным историческим источником. Можно провести параллели между многими культурно-хронологическими группами и соответствующими погребальными археологическими памятниками, чтобы добавить существенную информацию о той или иной культуре, полученную в результате раскопок. Некоторые периоды в этом районе пока не представлены с точки зрения археологии, кроме наскального искусства (вероятнее всего, артефакты еще не найдены).В этом случае петроглифы дают информацию неоценимого значения. Наиболее типичным примером является изображение так называемого «древнего слоя». Фигурки диких животных, а также загадочные антропоморфные фигуры и символические изображения лодок с «пассажирами» выполнены с большим реализмом и художественным мастерством. Датировка их неизвестна, установлено лишь, что не позднее III тысячелетия до н.э. Не исключено, что они относятся к каменному веку. Когда-то все прибрежные скалы сплошь были покрыты такими росписями.Не все из них сохранились до наших дней, но те, что уцелели, являются впечатляющим образцом художественного гения первобытного человека и дают нам ценные сведения, не подтвержденные до сих пор палеонтологическими источниками, об исчезнувших из района биоценозах. Некоторые животные в наскальных рисунках вымерли, а другие больше не обитали в этой местности. К первым относятся дикая лошадь (Equus ferus) и первобытный бык (Bos taurus primigenius). К последним относятся лось (Alces alces), благородный олень ( Cervus elaphus ), горный козёл ( Capra sibirica ), горный баран ( Ovis ammon ), медведь ( Ursus arctos ) и кабан. Sus scrofa ).Единственный вид, который до сих пор обитает на территории ареала – это косуля ( Capreolus pygargus ).
Наиболее типичным примером является изображение так называемого «древнего слоя». Фигурки диких животных, а также загадочные антропоморфные фигуры и символические изображения лодок с «пассажирами» выполнены с большим реализмом и художественным мастерством. Датировка их неизвестна, установлено лишь, что не позднее III тысячелетия до н.э. Не исключено, что они относятся к каменному веку. Когда-то все прибрежные скалы сплошь были покрыты такими росписями.Не все из них сохранились до наших дней, но те, что уцелели, являются впечатляющим образцом художественного гения первобытного человека и дают нам ценные сведения, не подтвержденные до сих пор палеонтологическими источниками, об исчезнувших из района биоценозах. Некоторые животные в наскальных рисунках вымерли, а другие больше не обитали в этой местности. К первым относятся дикая лошадь (Equus ferus) и первобытный бык (Bos taurus primigenius). К последним относятся лось (Alces alces), благородный олень ( Cervus elaphus ), горный козёл ( Capra sibirica ), горный баран ( Ovis ammon ), медведь ( Ursus arctos ) и кабан. Sus scrofa ).Единственный вид, который до сих пор обитает на территории ареала – это косуля ( Capreolus pygargus ).
Среди образов более поздних эпох есть такие, которые отражают влияние культур отдаленных территорий (Средней Азии, Китая и др.), прямые миграции и присутствие тех или иных этносов служат косвенным свидетельством исторических событий. На скалах встречаются изображения культурных артефактов, например, на уникальном предмете – плите из Кизанского района выбита серия изображений кинжалов и колесниц эпохи поздней бронзы.На нижних плитах горы «Сорок Зубьев» есть изображение котла и, вероятно, самое древнее изображение войлочной юрты — типичного жилища кочевников. Однако абсолютное большинство изображений имеет мифологические, эпические и религиозные сюжеты и служит источником для реконструкции мировоззрений разных эпох и народов.
Отдельного упоминания заслуживает серия изображений так называемого этнографического времени. Это рисунки коренного населения — хакасов, которые продолжали поддерживать тысячелетнюю традицию наскального искусства в Оглахтах вплоть до начала ХХ века. В Хакасии нет другого памятника с таким количеством хакасской живописи. Они очень информативны как для изучения верований и мифологии хакасов, так и реалий их быта, хозяйственного уклада и даже исторических событий. На сгибах и отдельных пластинах изображения лошадей (украшенных, с тамгами, «пятнистых», с «бубенчиками», оседланных, запряженных, с всадниками и без них и др.), верблюдов, оленей, собак, козлов. Многочисленные антропоморфные фигуры, одетые по-разному, символически, держащиеся за руки, образующие ряды из десятков фигур с луками, ружьями и т. д.; много изображений шаманов в одеяниях с бахромой, с барабанами и тампонами.Изображения тамги и других знаков, а также загадочные сложные абстрактные мотивы встречаются в большом количестве. Образы христианской церкви, самовара, телеги и т.п. свидетельствуют о проникновении русской культуры. Существенно, что некоторые сюжеты не могут быть объяснены с точки зрения имеющихся этнографических источников данных, то есть представляют собой уникальный источник для дальнейшего изучения традиционной культуры хакасов.
В Хакасии нет другого памятника с таким количеством хакасской живописи. Они очень информативны как для изучения верований и мифологии хакасов, так и реалий их быта, хозяйственного уклада и даже исторических событий. На сгибах и отдельных пластинах изображения лошадей (украшенных, с тамгами, «пятнистых», с «бубенчиками», оседланных, запряженных, с всадниками и без них и др.), верблюдов, оленей, собак, козлов. Многочисленные антропоморфные фигуры, одетые по-разному, символически, держащиеся за руки, образующие ряды из десятков фигур с луками, ружьями и т. д.; много изображений шаманов в одеяниях с бахромой, с барабанами и тампонами.Изображения тамги и других знаков, а также загадочные сложные абстрактные мотивы встречаются в большом количестве. Образы христианской церкви, самовара, телеги и т.п. свидетельствуют о проникновении русской культуры. Существенно, что некоторые сюжеты не могут быть объяснены с точки зрения имеющихся этнографических источников данных, то есть представляют собой уникальный источник для дальнейшего изучения традиционной культуры хакасов.
Таким образом, на номинируемой территории, несмотря на ее небольшие размеры, сосредоточено огромное количество разнообразных разновременных объектов наследия, которые как нельзя лучше характеризуют развитие культуры всего региона на протяжении тысячелетий.
границ | Нелинейный ответ на количество клеток, обнаруженный и исключенный из многолетних измерений трахеид сосны обыкновенной в Южной Сибири
Введение
Климат оказывает глубокое влияние на рост и диапазон распространения древесных пород, и, следовательно, более глубокое понимание взаимосвязи роста деревьев и климата будет способствовать их лучшему управлению и сохранению в условиях меняющихся климатических сценариев (Weiskopf et al., 2020; Upadhyay и другие., 2021). Это становится обязательным, особенно в таких районах, как Южная Сибирь, где леса, растущие вдоль засушливых лесных полос, уязвимы даже при небольшом изменении режима увлажнения (Xu et al., 2017). Дендроклиматические исследования позволяют нам получить важные эмпирические данные о долгосрочном влиянии климата на рост и развитие лесных деревьев. Классическая дендроклиматология определяет климатические факторы, ответственные за существенные вариации количественных характеристик годичных колец, и обеспечивает их реконструкцию по годичным кольцам в рамках обратной задачи (Fritts, 1976; Hughes et al., 2010; Швайнгрубер, 2012). Такие исследования были первоначально начаты с базовой характеристики, ширины годичных колец, как регистратора влияния климата на рост деревьев (Kapteyn, 1914; Douglass, 1920, 1925; Bailey, 1948; Glock, 1955), и продолжились изучением спектра переменные, а именно ширина зон ранней и поздней древесины, характеристики плотности древесины, изотопный и химический состав и, наконец, количественная анатомия древесины (КВА) вплоть до масштаба клеток (Fry, Chalk, 1957; Zahner et al., 1964). ; Хьюз и др., 2010; Эспер и др., 2016; Юнгквист и др., 2020). Классическая дендроклиматология предоставляет жизненно важную информацию о взаимодействии роста деревьев и климата, тогда как более точные объяснения реакции деревьев на изменения окружающей среды раскрываются в исследованиях ксилогенеза и возникающих в результате специфических черт древесины (De Micco et al.
Классическая дендроклиматология определяет климатические факторы, ответственные за существенные вариации количественных характеристик годичных колец, и обеспечивает их реконструкцию по годичным кольцам в рамках обратной задачи (Fritts, 1976; Hughes et al., 2010; Швайнгрубер, 2012). Такие исследования были первоначально начаты с базовой характеристики, ширины годичных колец, как регистратора влияния климата на рост деревьев (Kapteyn, 1914; Douglass, 1920, 1925; Bailey, 1948; Glock, 1955), и продолжились изучением спектра переменные, а именно ширина зон ранней и поздней древесины, характеристики плотности древесины, изотопный и химический состав и, наконец, количественная анатомия древесины (КВА) вплоть до масштаба клеток (Fry, Chalk, 1957; Zahner et al., 1964). ; Хьюз и др., 2010; Эспер и др., 2016; Юнгквист и др., 2020). Классическая дендроклиматология предоставляет жизненно важную информацию о взаимодействии роста деревьев и климата, тогда как более точные объяснения реакции деревьев на изменения окружающей среды раскрываются в исследованиях ксилогенеза и возникающих в результате специфических черт древесины (De Micco et al. , 2019).
, 2019).
Недавние усовершенствования методов раскроя древесины и технологический прогресс в измерении (полуавтоматическом и автоматическом) анатомических признаков клеток открыли новые возможности в дендроклиматологии (von Arx and Carrer, 2014; von Arx et al., 2016; Арзак и др., 2018 г.; Питерс и др., 2018 г.; Гебрегеоргис и др., 2021). Например, количественная анатомия отдельных древесно-кольцевых зон по трахеидограммам стала одним из перспективных подходов (Ваганов и др., 2006; Каррер и др., 2017; Мартин-Бенито и др., 2017), которые формируют дендроклиматологию высокое временное разрешение по шкале коротких интервалов в течение вегетационного периода (Pritzkow et al., 2014; Ziaco et al., 2016; Souto-Herrero et al., 2017; Pandey, 2021). Однако при использовании характеристик QWA в качестве новых дендроклиматических индикаторов были обнаружены статистические проблемы в связи с их возможным соответствием как климатическим переменным, так и между собой, что видно из многочисленных прямых наблюдений за сезонной кинетикой прироста годичных колец (Cuny, Rathgeber, 2016). ; Куни и др., 2019; Де Микко и др., 2019 г.; Виейра и др., 2020 г.; Стэнглер и др., 2021). У хвойных в развитии ксилемы наблюдаются три основных процесса (т. е. продукция, рост клеток за счет расширения и отложение вторичной клеточной стенки) последовательно для каждой трахеиды (Larson, 1994; Plomion et al., 2001; Fromm, 2013). Последовательный порядок образования клеток в радиальных рядах годичных колец в результате камбиальной активности в течение сезона и их созревания предоставляет информацию в тонком пространственно-временном масштабе для выделения внешнего сигнала в результирующих анатомических признаках (Castagneri et al. ., 2017, 2018; ср. Каррер и др., 2017). Однако возможные регуляторные связи между последовательными процессами ксилогенеза и степенью самостоятельности их реакций на внешние факторы во многом неизвестны и обсуждаются (Vaganov et al., 2011; Olano et al., 2012; Cuny et al., 2014; De Micco). и др., 2019). В некоторых исследованиях каждый этап ксилогенеза рассматривается как самостоятельный регистратор среды в пределах его продолжительности.
; Куни и др., 2019; Де Микко и др., 2019 г.; Виейра и др., 2020 г.; Стэнглер и др., 2021). У хвойных в развитии ксилемы наблюдаются три основных процесса (т. е. продукция, рост клеток за счет расширения и отложение вторичной клеточной стенки) последовательно для каждой трахеиды (Larson, 1994; Plomion et al., 2001; Fromm, 2013). Последовательный порядок образования клеток в радиальных рядах годичных колец в результате камбиальной активности в течение сезона и их созревания предоставляет информацию в тонком пространственно-временном масштабе для выделения внешнего сигнала в результирующих анатомических признаках (Castagneri et al. ., 2017, 2018; ср. Каррер и др., 2017). Однако возможные регуляторные связи между последовательными процессами ксилогенеза и степенью самостоятельности их реакций на внешние факторы во многом неизвестны и обсуждаются (Vaganov et al., 2011; Olano et al., 2012; Cuny et al., 2014; De Micco). и др., 2019). В некоторых исследованиях каждый этап ксилогенеза рассматривается как самостоятельный регистратор среды в пределах его продолжительности. Поскольку процесс дифференцировки трахеид начинается с камбиальной зоны (Larson, 1994; Plomion et al., 2001; Мияшима и др., 2013 г.; Рамос и Риган, 2018 г.; Aloni, 2021), в других работах деление клеток признается основным рецептором внешних воздействий, передающим полученный климатический сигнал на дальнейшие этапы дифференцировки клеток посредством внутренних обратных связей (Ваганов и др., 2011, 2020). Такой унаследованный эффект связывает величину и время действия климатических факторов во время деления трахеид с их последующим развитием через камбиальную зону. На его существование косвенно указывают устойчивые значимые связи, обнаруженные для морфометрических параметров одних и тех же трахеид или зон годичных колец между собой и с клеточной продукцией (Castagneri et al., 2017, 2018; Белокопытова и др., 2020; Пьерматтеи и др., 2020 г.; Жирнова и др., 2021). Поэтому рекомендуется учитывать наличие этого унаследованного эффекта при попытке использовать QWA в дендроклиматическом анализе и реконструкциях.
Поскольку процесс дифференцировки трахеид начинается с камбиальной зоны (Larson, 1994; Plomion et al., 2001; Мияшима и др., 2013 г.; Рамос и Риган, 2018 г.; Aloni, 2021), в других работах деление клеток признается основным рецептором внешних воздействий, передающим полученный климатический сигнал на дальнейшие этапы дифференцировки клеток посредством внутренних обратных связей (Ваганов и др., 2011, 2020). Такой унаследованный эффект связывает величину и время действия климатических факторов во время деления трахеид с их последующим развитием через камбиальную зону. На его существование косвенно указывают устойчивые значимые связи, обнаруженные для морфометрических параметров одних и тех же трахеид или зон годичных колец между собой и с клеточной продукцией (Castagneri et al., 2017, 2018; Белокопытова и др., 2020; Пьерматтеи и др., 2020 г.; Жирнова и др., 2021). Поэтому рекомендуется учитывать наличие этого унаследованного эффекта при попытке использовать QWA в дендроклиматическом анализе и реконструкциях. Кроме того, его удаление из временных рядов анатомических признаков трахеид может быть выполнено с использованием методов, аналогичных тем, которые применяются для удаления долгосрочных трендов из измерений ширины годичных колец при их стандартизации (Cook, Kairiukstis, 1990). В частности, предлагается моделирование зависимостей анатомических признаков от числа клеток в кольце (как характеристика камбиальной активности) с последующей индексацией анатомических хронологий в виде отношения к полученным модельным значениям.Мы предполагаем, что такая индексация может помочь отделить климатические сигналы, непосредственно воспринимаемые анатомическими признаками, от наследия камбиальной активности, отраженного в числе клеток. Внутренние отношения между признаками древесины лучше всего можно описать с помощью набора данных, охватывающих всю или, по крайней мере, большую часть жизни дерева, т. е. максимизируя длину анатомической хронологии, чтобы уменьшить погрешность, связанную с возрастом. Однако из-за трудоемкости и ресурсоемкости исследования в этой области в основном оперируют наборами анатомических данных продолжительностью в несколько десятилетий или редко достигающих столетия (Yasue et al.
Кроме того, его удаление из временных рядов анатомических признаков трахеид может быть выполнено с использованием методов, аналогичных тем, которые применяются для удаления долгосрочных трендов из измерений ширины годичных колец при их стандартизации (Cook, Kairiukstis, 1990). В частности, предлагается моделирование зависимостей анатомических признаков от числа клеток в кольце (как характеристика камбиальной активности) с последующей индексацией анатомических хронологий в виде отношения к полученным модельным значениям.Мы предполагаем, что такая индексация может помочь отделить климатические сигналы, непосредственно воспринимаемые анатомическими признаками, от наследия камбиальной активности, отраженного в числе клеток. Внутренние отношения между признаками древесины лучше всего можно описать с помощью набора данных, охватывающих всю или, по крайней мере, большую часть жизни дерева, т. е. максимизируя длину анатомической хронологии, чтобы уменьшить погрешность, связанную с возрастом. Однако из-за трудоемкости и ресурсоемкости исследования в этой области в основном оперируют наборами анатомических данных продолжительностью в несколько десятилетий или редко достигающих столетия (Yasue et al. , 2000; Кирдянов и др., 2003; Панюшкина и др., 2003; Кастаньери и др., 2017). В этом исследовании была разработана долгосрочная хронология (более 200 лет) анатомических признаков древесины с основным акцентом на оценку взаимосвязей между измеренными характеристиками древесины. Основное внимание в исследовании уделялось выявлению любых существующих взаимосвязей между числом клеток в кольце и характеристиками трахеидограмм, соответствующих двум последующим фазам дифференцировки, а также выявлению и количественной оценке формы этих взаимосвязей (линейной или нелинейной).Это исследование было проведено со следующими основными целями: (1) определить, зависят ли радиальный размер ячейки и толщина стенки от количества клеток в радиальном ряду кольца дерева (что указывает на вышеупомянутый унаследованный эффект), (2) чтобы выразить эти зависимости аналитически, ( 3) разработать индексированные анатомические хронологии с устранением унаследованного эффекта путем деления фактических значений на смоделированные и (4) установить разделение климатических реакций в этих «уточненных» анатомических хронологиях.
, 2000; Кирдянов и др., 2003; Панюшкина и др., 2003; Кастаньери и др., 2017). В этом исследовании была разработана долгосрочная хронология (более 200 лет) анатомических признаков древесины с основным акцентом на оценку взаимосвязей между измеренными характеристиками древесины. Основное внимание в исследовании уделялось выявлению любых существующих взаимосвязей между числом клеток в кольце и характеристиками трахеидограмм, соответствующих двум последующим фазам дифференцировки, а также выявлению и количественной оценке формы этих взаимосвязей (линейной или нелинейной).Это исследование было проведено со следующими основными целями: (1) определить, зависят ли радиальный размер ячейки и толщина стенки от количества клеток в радиальном ряду кольца дерева (что указывает на вышеупомянутый унаследованный эффект), (2) чтобы выразить эти зависимости аналитически, ( 3) разработать индексированные анатомические хронологии с устранением унаследованного эффекта путем деления фактических значений на смоделированные и (4) установить разделение климатических реакций в этих «уточненных» анатомических хронологиях.
Материалы и методы
Зона исследования
Исследования проводились в восточной части Батеньского хребта Кузнецкого Алатау на юге Сибири (рис. 1А,Б). Высота хребта колеблется от 500 м над ур. в предгорьях до 1200 м над ур. на вершинах гор в центральной части. Смешанный лес из лиственницы сибирской ( Larix sibirica Ledeb.), сосны обыкновенной ( Pinus sylvestris L.) и березы повислой ( Betula pendula Roth.) в разных пропорциях занимает большую часть хребта, но на более сухих участках южных и юго-восточных склонов, особенно в предгорьях, встречаются редколесья, чередующиеся со степной растительностью.
Рисунок 1 . Учебная зона. Территориальное расположение пункта отбора проб (БГД; треугольник) и метеостанции Шира (ШР; звездочка) в северной части хребта Батень горной системы Кузнецкий Алатау (А) . Расположение исследуемой территории (прямоугольник) в Сибири, Россия (Б) ; цветом отмечен административный район Республики Хакасия. Климатическая диаграмма среднемесячных температур (1966-2008 гг.; линия) и осадков (1936-2008 гг., бары) на ст. Шира (С) . Фоновые карты: спутниковые данные © Google, 2020; схематическая карта является фрагментом Карты Хакасии в пределах России (CC BY-SA 4.0).
Климатическая диаграмма среднемесячных температур (1966-2008 гг.; линия) и осадков (1936-2008 гг., бары) на ст. Шира (С) . Фоновые карты: спутниковые данные © Google, 2020; схематическая карта является фрагментом Карты Хакасии в пределах России (CC BY-SA 4.0).
Климат района резко континентальный (Алисов, 1956; рис. 1В), характеризующийся большими сезонными и суточными перепадами температур. По данным ближайшей метеостанции Шира (54,50° с.ш., 59,93° в.д., 475 м над ур.с.л.), среднегодовая температура района исследований составляет ~+0,9°С при годовом количестве осадков около 300 мм. Как правило, в горных условиях более низкие температуры и более обильные осадки по сравнению с равнинными районами, где расположена станция Шира. Осадки в районе характеризовались неравномерным годовым распределением, при этом большая часть осадков (> 80%) выпадала в сезон положительных температур, с наибольшим количеством осадков в июле. Средний безморозный период положительных минимальных температур длится с мая по сентябрь, когда среднесуточные температуры фиксируются выше +5°С. Дендроклиматический анализ выполнен с использованием суточных климатических рядов осадков (1936–2008 гг.) и температуры (1966–2008 гг.) на ст. Шира. Климатические данные получены из Всероссийского научно-исследовательского института гидрометеорологической информации, Мировой центр данных (ВНИИГМИ-МЦД; http://meteo.ru/data).
Дендроклиматический анализ выполнен с использованием суточных климатических рядов осадков (1936–2008 гг.) и температуры (1966–2008 гг.) на ст. Шира. Климатические данные получены из Всероссийского научно-исследовательского института гидрометеорологической информации, Мировой центр данных (ВНИИГМИ-МЦД; http://meteo.ru/data).
Сбор проб, измерение и обработка данных годичных колец
Образцы древесины сосны обыкновенной (керны) отобраны в окрестностях села Боград, расположенного на северо-восточной оконечности хребта (БГД; 54.20–54,27° с.ш., 90,68–90,92° в.д., 500–650 м над ур. Первоначальный отбор проб был проведен в 1989 г., который впоследствии был повторен в течение 2012–2018 гг. Всего за весь период было отобрано 86 деревьев для выявления самых старых сосен в районе. Образцы керна отбирали со здоровых неповрежденных взрослых деревьев верхнего яруса полога в пределах древостоя (высота деревьев ~10–12 м) по склонам южной, юго-западной и юго-восточной экспозиций, где повышенная инсоляция повышает чувствительность деревьев к влаге. дефицит и усиливает климатический сигнал.Отбор проб производили приращенным буром на высоте груди (~1,3 м) перпендикулярно склону, т. е. преимущественно в западно-восточном направлении. Собранные образцы были доставлены в лабораторию и подготовлены для измерений в соответствии со стандартными протоколами, изложенными Куком и Кайрюкстисом (1990). Ширину годичных колец деревьев ( TRW ) серии каждого образца измеряли с использованием системы LINTAB-5 и программы TSAP-win (Rinn, 2003). Точность перекрестного датирования ряда была проверена с использованием программы COFECHA (Holmes, 1983).Наконец, стандартизированная локальная хронология TRW была разработана с использованием компьютерной программы ARSTAN (Cook and Krusic, 2005). Во время процедуры стандартизации долгосрочные тренды в отдельных сериях TRW описывались 67% кубическим сглаживающим сплайном с 50% отсечкой частотной характеристики (Cook and Peters, 1981).
дефицит и усиливает климатический сигнал.Отбор проб производили приращенным буром на высоте груди (~1,3 м) перпендикулярно склону, т. е. преимущественно в западно-восточном направлении. Собранные образцы были доставлены в лабораторию и подготовлены для измерений в соответствии со стандартными протоколами, изложенными Куком и Кайрюкстисом (1990). Ширину годичных колец деревьев ( TRW ) серии каждого образца измеряли с использованием системы LINTAB-5 и программы TSAP-win (Rinn, 2003). Точность перекрестного датирования ряда была проверена с использованием программы COFECHA (Holmes, 1983).Наконец, стандартизированная локальная хронология TRW была разработана с использованием компьютерной программы ARSTAN (Cook and Krusic, 2005). Во время процедуры стандартизации долгосрочные тренды в отдельных сериях TRW описывались 67% кубическим сглаживающим сплайном с 50% отсечкой частотной характеристики (Cook and Peters, 1981).
В 2019 году для анатомических измерений было отобрано семь кернов (по одному керну на дерево) из полной коллекции образцов. Ядра были отобраны на основе максимального камбиального возраста (количество годичных колец от коры до сердцевины), высоких корреляций TRW с местной хронологией (дополнительный рисунок 1) и их целостности.На ротационном микротоме (Microm HM 340 E; Thermo Fisher Scientific, США) были получены поперечные срезы толщиной 12–14 мкм из выбранных ядер деревьев без учета участков ювенильной древесины (самые внутренние 10–15 колец, заметно различающихся по анатомическому строению). ), чтобы свести к минимуму связанные с высотой тенденции в деревянной гидравлической архитектуре (Mencuccini et al., 1997), если они присутствовали. Поперечные срезы окрашивали сафранином и астровым синим, обезвоживали в этаноле возрастающей концентрации, промывали ксилолом и помещали в канадский бальзам на предметные стекла.
Ядра были отобраны на основе максимального камбиального возраста (количество годичных колец от коры до сердцевины), высоких корреляций TRW с местной хронологией (дополнительный рисунок 1) и их целостности.На ротационном микротоме (Microm HM 340 E; Thermo Fisher Scientific, США) были получены поперечные срезы толщиной 12–14 мкм из выбранных ядер деревьев без учета участков ювенильной древесины (самые внутренние 10–15 колец, заметно различающихся по анатомическому строению). ), чтобы свести к минимуму связанные с высотой тенденции в деревянной гидравлической архитектуре (Mencuccini et al., 1997), если они присутствовали. Поперечные срезы окрашивали сафранином и астровым синим, обезвоживали в этаноле возрастающей концентрации, промывали ксилолом и помещали в канадский бальзам на предметные стекла.
Микрофотографии поперечных срезов получены с помощью цифровой камеры (ProgRes Gryphax Subra, Jenoptik GmbH, Германия), установленной на оптическом микроскопе с 200-кратным увеличением (BX43, Olympus, Япония). Среди рассматриваемых годичных колец наблюдались многочисленные внутригодовые колебания плотности (IADF) в различных положениях внутри кольца. Некоторые из узких колец показали более тонкие клеточные стенки в поздней древесине (светлые кольца). Морозные кольца и синие кольца не наблюдались. Анатомические признаки всех колец в отобранных образцах измеряли с помощью полуавтоматической программы «Линейка 2».01 (Силкин, 2010). Границы годичных колец были определены пользователем вручную. Анатомические признаки древесины, а именно число клеток ( N ), их радиальный диаметр ( D ) и толщину клеточных стенок ( CWT ), измеряли (Ваганов и др., 1985; Ларсон, 1994) для пять радиальных рядов клеток в каждом кольце (Seo et al., 2014). Для получения правильных средних оценок анатомических признаков в пределах каждого конкретного кольца внутрисезонные серии D и CWT (трахеидограммы) нормализовали путем их сжатия/расширения на такое же количество клеток, как усредненное по пяти измеренным радиальным рядам (Ваганов, 1990).
Среди рассматриваемых годичных колец наблюдались многочисленные внутригодовые колебания плотности (IADF) в различных положениях внутри кольца. Некоторые из узких колец показали более тонкие клеточные стенки в поздней древесине (светлые кольца). Морозные кольца и синие кольца не наблюдались. Анатомические признаки всех колец в отобранных образцах измеряли с помощью полуавтоматической программы «Линейка 2».01 (Силкин, 2010). Границы годичных колец были определены пользователем вручную. Анатомические признаки древесины, а именно число клеток ( N ), их радиальный диаметр ( D ) и толщину клеточных стенок ( CWT ), измеряли (Ваганов и др., 1985; Ларсон, 1994) для пять радиальных рядов клеток в каждом кольце (Seo et al., 2014). Для получения правильных средних оценок анатомических признаков в пределах каждого конкретного кольца внутрисезонные серии D и CWT (трахеидограммы) нормализовали путем их сжатия/расширения на такое же количество клеток, как усредненное по пяти измеренным радиальным рядам (Ваганов, 1990). ).В связи с трудоемкостью и сложностью выделения зон ранней и поздней древесины в многочисленных кольцах с ИАПД предпочтение было отдано расчету характеристик клеток, не зависящих от достоверности критериев, используемых для разделения зон. Характеристики ячейки, а именно максимальный ( D max ) и средний ( D средний ) радиальный диаметр, а также максимальный ( CWT max ) и средний ( CWT средний ) толщина стенки , рассчитывали для каждого кольца по нормализованным усредненным трахеидограммам (рис. 2).Надежность этих клеточных характеристик была проверена путем сравнения средних значений с медианой и максимальных значений с 90-м процентилем для небольшой подвыборки случайно выбранных годичных колец как в отдельных радиальных рядах трахеид, так и в средних трахеидограммах (дополнительный рисунок 2). Средние значения D и CWT интегрируют сигнал окружающей среды на соответствующих стадиях развития клеток для всех клеток древесного кольца таким же образом, как N ассимилирует сигнал окружающей среды во время клеточного деления.
).В связи с трудоемкостью и сложностью выделения зон ранней и поздней древесины в многочисленных кольцах с ИАПД предпочтение было отдано расчету характеристик клеток, не зависящих от достоверности критериев, используемых для разделения зон. Характеристики ячейки, а именно максимальный ( D max ) и средний ( D средний ) радиальный диаметр, а также максимальный ( CWT max ) и средний ( CWT средний ) толщина стенки , рассчитывали для каждого кольца по нормализованным усредненным трахеидограммам (рис. 2).Надежность этих клеточных характеристик была проверена путем сравнения средних значений с медианой и максимальных значений с 90-м процентилем для небольшой подвыборки случайно выбранных годичных колец как в отдельных радиальных рядах трахеид, так и в средних трахеидограммах (дополнительный рисунок 2). Средние значения D и CWT интегрируют сигнал окружающей среды на соответствующих стадиях развития клеток для всех клеток древесного кольца таким же образом, как N ассимилирует сигнал окружающей среды во время клеточного деления. Максимальные значения D и CWT тесно связаны со средним значением D в ранней древесине и средним значением CWT в поздней древесине соответственно. Предыдущие исследования указывали на то, что для хвойных в районе исследований эти зависимости носят устойчиво линейный характер (Жирнова и др., 2021; неопубликованные данные для сосны обыкновенной в различных местообитаниях), что, вероятно, связано с подавлением стохастических выбросов путем усреднения измерений пяти радиальных рядов. в каждом кольце. Корреляции максимальных и средних значений высокие, 0.89–0,96 для отдельных деревьев и 0,95 в среднем для D и 0,81–0,94 и 0,74 для CWT , что означает 66–92% общей дисперсии. Однако мы ожидаем еще более высоких корреляционных связей между D max и ранней древесиной D, CWT max и поздней древесиной CWT (для сравнения, для сосны обыкновенной вблизи верхней границы леса в исследуемом районе наблюдались следующие корреляции).
Максимальные значения D и CWT тесно связаны со средним значением D в ранней древесине и средним значением CWT в поздней древесине соответственно. Предыдущие исследования указывали на то, что для хвойных в районе исследований эти зависимости носят устойчиво линейный характер (Жирнова и др., 2021; неопубликованные данные для сосны обыкновенной в различных местообитаниях), что, вероятно, связано с подавлением стохастических выбросов путем усреднения измерений пяти радиальных рядов. в каждом кольце. Корреляции максимальных и средних значений высокие, 0.89–0,96 для отдельных деревьев и 0,95 в среднем для D и 0,81–0,94 и 0,74 для CWT , что означает 66–92% общей дисперсии. Однако мы ожидаем еще более высоких корреляционных связей между D max и ранней древесиной D, CWT max и поздней древесиной CWT (для сравнения, для сосны обыкновенной вблизи верхней границы леса в исследуемом районе наблюдались следующие корреляции). : r ( D макс. , D среднее ) = 0.85, R ( D ( D MAX , D EW ) = 0,92, R ( CWT MAX , CWT означает ) = 0,89, R ( CWT max , CWT lw ) = 0,97; Жирнова и др., 2021). Следовательно, D max и CWT max можно интерпретировать как легко выводимые замены зональных характеристик, и они должны регистрировать экологический сигнал в более краткосрочных интервалах, чем D означает и CWT . означают , несмотря на сильную связь между ними.
: r ( D макс. , D среднее ) = 0.85, R ( D ( D MAX , D EW ) = 0,92, R ( CWT MAX , CWT означает ) = 0,89, R ( CWT max , CWT lw ) = 0,97; Жирнова и др., 2021). Следовательно, D max и CWT max можно интерпретировать как легко выводимые замены зональных характеристик, и они должны регистрировать экологический сигнал в более краткосрочных интервалах, чем D означает и CWT . означают , несмотря на сильную связь между ними.
Рисунок 2 . Методика расчета максимальных и средних значений признаков клеток: усредненные трахеидограммы радиального диаметра, D и толщины клеточной стенки, CWT (A) ; микрофотография годичных колец, образовавшихся в 1903 году на дереве № 6, используемая в качестве примера для расчета (B) .
Долгосрочные хронологии этих четырех клеточных характеристик были разработаны для каждого отдельного дерева и, наконец, усреднены для всех семи деревьев, чтобы получить локальные хронологии.Исключение самых внутренних колец свело к минимуму долгосрочные тенденции, связанные с размером деревьев, в анатомических сериях, поскольку не наблюдалось значительных линейных тенденций при p <0,1 ни в общей серии, ни в самый ранний 50-летний период. Поэтому детрендирование анатомических хронологий не проводилось. Отдельные серии TRW были рассчитаны как сумма D . Они использовались для кросс-датирования серии анатомических измерений с предварительными измерениями LINTAB тех же кернов.Подсерии анатомически полученных TRW для каждой части керна были отдельно перекрестно датированы с полными сериями TRW , измеренными на LINTAB ( r > 0,9), а затем все датированные подсерии были объединены в одну серию. Кроме того, эти серии TRW были перекрестно датированы с местной хронологией TRW ( r > 0,7), полученной из кернов всех 86 деревьев, собранных на месте отбора проб (дополнительный рисунок 1). Некоторые кольца в полученной таким образом хронологии отсутствовали как недостающие кольца (год без роста дерева из-за неблагоприятных условий).В ряде случаев кольца на границах кусков керна механически повреждались при резке и поэтому отбраковывались на этапе измерения.
Эффект наследия камбиальной зоны в анатомических признаках был оценен для вышеупомянутых генерализованных характеристик трахеид. Номер клетки N интерпретировался здесь как результирующая характеристика камбиальной активности, агрегированная за весь вегетационный сезон. Точно так же средние измерения трахеид использовались в качестве агрегированных результирующих характеристик последующих стадий ксилогенеза.Хотя образование и дифференцировка наиболее крупных клеток и наиболее толстостенных клеток в годичных кольцах деревьев приходится ограничивать определенными меньшими временными интервалами в течение вегетационного сезона, подробные сведения о продукции клеток в эти интервалы отсутствуют; таким образом, эти максимальные значения также сравнивали с общим числом клеток. Зависимости были аналитически описаны нелинейными регрессионными моделями D MAX ( N ), D Среднее ( N ), CWT MAX ( N ) и CWT означает ( N ) с использованием уравнений и числовых выражений, направленных на лучшую статистическую пригодность моделей (максимизация R 2 ) и на простоту экологической интерпретации.Модели построены с использованием унифицированной формы уравнения, а числовые члены разрабатывались отдельно для каждого рассматриваемого дерева в связи с ожидаемыми различиями в требованиях к водопроводности и механической прочности ксилемы в силу аллометрии деревьев, генетической предрасположенности и микроусловий (Mencuccini и др., 1997; Анфодилло и др., 2012, 2013). Устранение эффекта наследия было выполнено путем расчета индексированных рядов (отношение фактических значений к смоделированным) для каждого дерева и их усреднения для получения локальной хронологии.
Зависимости были аналитически описаны нелинейными регрессионными моделями D MAX ( N ), D Среднее ( N ), CWT MAX ( N ) и CWT означает ( N ) с использованием уравнений и числовых выражений, направленных на лучшую статистическую пригодность моделей (максимизация R 2 ) и на простоту экологической интерпретации.Модели построены с использованием унифицированной формы уравнения, а числовые члены разрабатывались отдельно для каждого рассматриваемого дерева в связи с ожидаемыми различиями в требованиях к водопроводности и механической прочности ксилемы в силу аллометрии деревьев, генетической предрасположенности и микроусловий (Mencuccini и др., 1997; Анфодилло и др., 2012, 2013). Устранение эффекта наследия было выполнено путем расчета индексированных рядов (отношение фактических значений к смоделированным) для каждого дерева и их усреднения для получения локальной хронологии. Этот подход дал лучшие результаты дальнейшего анализа (более близкое к нормальному распределение индексированных значений, более высокий общий сигнал и климатические корреляции для локальных хронологий) по сравнению с вычитанием смоделированных значений из фактических (данные не представлены).
Этот подход дал лучшие результаты дальнейшего анализа (более близкое к нормальному распределение индексированных значений, более высокий общий сигнал и климатические корреляции для локальных хронологий) по сравнению с вычитанием смоделированных значений из фактических (данные не представлены).
Климатический отклик осредненных местных хронологий TRW и индексированных анатомических параметров оценивался в виде корреляций со скользящими рядами (21-дневное окно, 1-дневный шаг внутри сезона) температуры и осадков, которые рассчитывались по суточным данным из Станция Шира.Анализ проводился для всего периода охвата климатическими данными и для скользящих 30-летних интервалов (шаг 1 год) для оценки временной устойчивости отклика.
Результаты
Продолжительность отдельных серий анатомических измерений варьировалась от 150 до 280 лет, охватывающих период 1739–2018 гг. Н. Э. (Дополнительный рисунок 1). Классический параметр ширины годичных колец показал линейную зависимость от количества клеток с помощью моделей линейной регрессии TRW ( N ) (рис. 3).Зависимость была почти прямо пропорциональной, о чем свидетельствует значение постоянного члена, не превышающее 120 мкм и являющееся недостоверным при p < 0,05. Наиболее широкие годичные кольца, содержащие наибольшее количество клеток, наблюдались у дерева № 3 с камбиальным возрастом 222 года, тогда как самый старый образец (дерево № 4, возраст 280 лет) показал наименьшую изменчивость ширины годичных колец и количества клеток. (Дополнительная таблица 1). Тем не менее значимых связей между камбиальным возрастом деревьев и диапазоном числа клеток в кольце не обнаружено.Для всех образцов коэффициент детерминации был близок к максимальному (0,95–0,98) и свидетельствовал о высокой достоверности полученных моделей при их строгой близости к линейной зависимости. Визуальное сравнение графиков, суммирующих информацию по всем смоделированным деревьям, показало почти линейность TRW (N) и в локальном масштабе. Большинство колец (95%) были обнаружены в диапазоне TRW < 1400 мкм и N < 45.
3).Зависимость была почти прямо пропорциональной, о чем свидетельствует значение постоянного члена, не превышающее 120 мкм и являющееся недостоверным при p < 0,05. Наиболее широкие годичные кольца, содержащие наибольшее количество клеток, наблюдались у дерева № 3 с камбиальным возрастом 222 года, тогда как самый старый образец (дерево № 4, возраст 280 лет) показал наименьшую изменчивость ширины годичных колец и количества клеток. (Дополнительная таблица 1). Тем не менее значимых связей между камбиальным возрастом деревьев и диапазоном числа клеток в кольце не обнаружено.Для всех образцов коэффициент детерминации был близок к максимальному (0,95–0,98) и свидетельствовал о высокой достоверности полученных моделей при их строгой близости к линейной зависимости. Визуальное сравнение графиков, суммирующих информацию по всем смоделированным деревьям, показало почти линейность TRW (N) и в локальном масштабе. Большинство колец (95%) были обнаружены в диапазоне TRW < 1400 мкм и N < 45.
Рисунок 3 .Зависимости ширины годичных колец от номера ячейки, TRW ( N ), для каждого исследуемого дерева и от масштаба пробной площадки (общая выборка). Маркеры (кружки) представляют фактические кольца деревьев, а линия представляет собой линейную функцию отношения (уравнение и коэффициент детерминации R 2 показаны на каждом графике).
Наследственный эффект производства клеток в QWA характеризовался зависимостью характеристик трахеид от количества клеток и хорошо описывался отрицательной экспоненциальной функцией (рис. 4, дополнительная рис. 3).Быстрое увеличение как средних, так и максимальных значений D и CWT на годичное дерево наблюдалось в узких кольцах синхронно с увеличением N , но постепенно переходило к стабильным значениям характеристик клеток в более широких кольцах с большими N . После выбора численных параметров для всех экспоненциальных моделей (таблица 1) корреляция между модельными значениями и фактическими измерениями составила от 0,53 до 0,86.
Рисунок 4 . Зависимости признаков трахеид от числа клеток, N , для максимального и среднего значений радиального диаметра D max ( N ) и D среднее ( N ) 1 A 9019 клеток толщина стенки CWT max ( N ) и CWT означает ( N ) (B) (на примере дерева №3).Маркеры точечных диаграмм (кружки) представляют собой годичные кольца деревьев; линии представляют экспоненциальные функции отношений. Данные о максимальных значениях признаков клеток ( D max , CWT max ) представлены черным цветом; серым цветом представлены данные о средних значениях клеточных признаков ( D среднее , CWT среднее ).
Таблица 1 . Численные параметры зависимостей размеров трахеид (радиальный диаметр клетки D и толщина клеточной стенки CWT ) от номера клетки N , выраженные в виде функций D = Dmin+(Das-Dmin)·(1-ea·N), CWT= CWTMIN + (CWTAS-CWTMIN) · (1-EA · N), установленные для максимального ( D ,
5 MAX , CWT MAX ) и среднее значение ( D означает , CWT Среднее ) Значения на ринге.
Индексы для каждого дерева были рассчитаны как отношение фактических измерений к соответствующим значениям модели, подход, аналогичный индексированию TRW в классической дендрохронологии. Индексированные значения имеют небольшую асимметрию и эксцесс, а также аналогичные значения медианы и среднего арифметического для большинства характеристик и отдельных деревьев (дополнительный рисунок 4, дополнительная таблица 2). Наконец, локальные хронологии рассматриваемых анатомических признаков были рассчитаны путем усреднения индивидуальных индексированных серий.Локальные хронологии охватывают период 279 лет, тогда как глубина выборки не менее трех деревьев получена за 211 лет (1807–2018 гг.), а пять деревьев охватывают период 185 лет (1834–2018 гг.) (рис. 5). Хронология характеристик CWT была стабильной во времени, в то время как индексированные хронологии как D max , так и D означают, что демонстрируют медленно возрастающие тенденции. Для периода 1807–2018 гг. наклоны этих трендов составляли около 0,001 отн. ед. в год с коэффициентами детерминации R 2 = 0.57 и R 2 = 0,48 при p < 0,01 для хронологий D max и D среднее соответственно. Тем не менее, для периодов дендроклиматического анализа R 2 < 0,2 наблюдалось для линейных многолетних трендов во всех четырех хронологиях, причем многолетняя вариация была значительно меньше, чем межгодовые колебания.
Для периода 1807–2018 гг. наклоны этих трендов составляли около 0,001 отн. ед. в год с коэффициентами детерминации R 2 = 0.57 и R 2 = 0,48 при p < 0,01 для хронологий D max и D среднее соответственно. Тем не менее, для периодов дендроклиматического анализа R 2 < 0,2 наблюдалось для линейных многолетних трендов во всех четырех хронологиях, причем многолетняя вариация была значительно меньше, чем межгодовые колебания.
Рисунок 5 . Локальные индексированные хронологии анатомических характеристик сосны обыкновенной: максимальное ( D max ) и среднее ( D среднее ) значения радиального диаметра клеток, максимальное ( CWT max ) и среднее ( CWT означают ) значения толщины клеточной стенки в кольце дерева.Линии представляют локальные средние хронологии; заштрихованные области представляют собой стандартные отклонения для каждого календарного года. На нижней панели показана динамика глубины выборки (количество отдельных серий) с учетом отсутствующих и выброшенных колец.
На нижней панели показана динамика глубины выборки (количество отдельных серий) с учетом отсутствующих и выброшенных колец.
Было отмечено, что исходные средние межсерийные корреляции (0,36–0,40) между анатомическими хронологиями семи отдельных деревьев существенно уменьшились после индексации ( D , 0,26–0,30; CWT , 0,11–0,13).Кроме того, средние корреляции отдельных серий с соответствующими локальными хронологиями были зарегистрированы как 0,67–0,70 до индексации для всех характеристик клеток, 0,64–0,66 для индексированного D и 0,51–0,55 для индексированного CWT .
Взаимосвязь между ростом деревьев и климатом для местных индексированных хронологий была проанализирована с использованием ежедневных данных о температуре (1966–2008 гг.) и осадках (1936–2008 гг.), обобщенных с окном в 21 день. Между локальными индексированными хронологиями и климатическими переменными наблюдались значительные взаимосвязи в течение сезона с 90 по 270 день года, т. е.э., 30 марта–26 сентября (рис. 6). TRW сосны обыкновенной положительно коррелирует с осадками и отрицательно с температурами с начала мая до середины июня. Напротив, для диаметра клеток наблюдалась иная картина ( D макс. и D среднее значение ), где влажные условия в период с мая по июнь благоприятствовали образованию крупных клеток, а температуры в этот период демонстрировали значительная отрицательная связь с D означает .С середины июня до конца июля выявлено негативное влияние обеих климатических переменных на размеры клеток, которое было более выражено в случае D сред . При этом CWT выявил зависимость от осадков с середины июня по начало августа, тогда как отрицательная связь температуры воздуха была более выражена с CWT max и наблюдалась в течение июля–августа. Тем не менее, положительная связь между CWT средним значением и температурами в мае добавила интересный факт в случае взаимосвязи роста деревьев и климата сосны обыкновенной.
е.э., 30 марта–26 сентября (рис. 6). TRW сосны обыкновенной положительно коррелирует с осадками и отрицательно с температурами с начала мая до середины июня. Напротив, для диаметра клеток наблюдалась иная картина ( D макс. и D среднее значение ), где влажные условия в период с мая по июнь благоприятствовали образованию крупных клеток, а температуры в этот период демонстрировали значительная отрицательная связь с D означает .С середины июня до конца июля выявлено негативное влияние обеих климатических переменных на размеры клеток, которое было более выражено в случае D сред . При этом CWT выявил зависимость от осадков с середины июня по начало августа, тогда как отрицательная связь температуры воздуха была более выражена с CWT max и наблюдалась в течение июля–августа. Тем не менее, положительная связь между CWT средним значением и температурами в мае добавила интересный факт в случае взаимосвязи роста деревьев и климата сосны обыкновенной.
Рисунок 6 . Климатическая реакция характеристик годичных колец сосны обыкновенной: ширина годичных колец ( TRW ), максимальное ( D max ) и среднее ( D среднее ) значения радиального диаметра клеток и максимальное ( CWT max ) и среднее ( CWT среднее ) значения толщины клеточной стенки в кольце дерева. На графиках показаны корреляции проиндексированных локальных хронологий с скользящими рядами температуры (красные линии; 1966–2008 гг.) и осадков (синие области; 1936–2008 гг.), рассчитанными по ежедневным данным с 21-дневным окном и 1-дневным шагом.Горизонтальные штриховые линии показывают уровень значимости p = 0,05 для корреляций с температурой (короткие штрихи) и осадками (длинные штрихи).
Анализ временной стабильности климатической реакции сосны обыкновенной Хронология TRW (дополнительный рисунок 5) показал снижение интенсивности реакции. Напротив, как максимальные, так и средние анатомические признаки древесных колец регистрировали постепенные сдвиги в интервалах значимого климатического сигнала в пределах сезона, но его интенсивность оставалась стабильной.И D max , и D mean показали максимальную положительную реакцию на осадки в более ранние сроки летнего сезона в последнее время, тогда как период их отрицательной реакции на температуру и осадки оставался фиксированным во второй половине лета. Напротив, CWT демонстрировал положительную реакцию на влажность в течение длительного периода (дата начала значимых корреляций стабильно совпадает с летним солнцестоянием, сочетаясь с более поздним окончанием реакции в последние десятилетия) со сдвигом отрицательной реакции температуре до более поздних сроков.
Напротив, как максимальные, так и средние анатомические признаки древесных колец регистрировали постепенные сдвиги в интервалах значимого климатического сигнала в пределах сезона, но его интенсивность оставалась стабильной.И D max , и D mean показали максимальную положительную реакцию на осадки в более ранние сроки летнего сезона в последнее время, тогда как период их отрицательной реакции на температуру и осадки оставался фиксированным во второй половине лета. Напротив, CWT демонстрировал положительную реакцию на влажность в течение длительного периода (дата начала значимых корреляций стабильно совпадает с летним солнцестоянием, сочетаясь с более поздним окончанием реакции в последние десятилетия) со сдвигом отрицательной реакции температуре до более поздних сроков.
Обсуждение
Наследие образования клеток в анатомических признаках годичных колец
Ширина древесных колец как сумма радиальных диаметров клеток усваивает результаты двух последовательных стадий дифференцировки трахеид, а именно деления клеток в камбиальной зоне и последующего их роста путем расширения в течение сезона роста (Larson, 1994; Fromm , 2013; Ратгебер и др. , 2016). Таким образом, любые отклонения функции TRW (N) от функции прямой пропорциональности (т.е., линейная функция с нулевым постоянным членом) были вызваны изменчивостью среднего D в годичном кольце. Эта изменчивость складывается из нескольких компонентов: (i) возрастных трендов ювенильной древесины (Высоцкая, Ваганов, 1989; Lei et al., 1996; Anfodillo et al., 2013), которые не учитываются в рассматриваемом наборе данных, (ii) унаследованный эффект, описываемый экспоненциальной функцией D ( N ), (iii) влияние текущих климатических колебаний во время расширения ячейки и (iv) стохастический компонент.Следовательно, функция TRW (N) имеет небольшую кривизну в области узких колец (рис. 3). Тем не менее, очень высокие коэффициенты детерминации, полученные для линейных моделей, свидетельствовали о высокой стабильности среднего размера клеток, образующихся у основания ствола дерева, на протяжении большей части жизни дерева. Напротив, близость этой зависимости утвердила TRW как параметр, интегрирующий влияние климатических колебаний преимущественно в период камбиальной деятельности и до ее начала (через состояние дерева в момент активизации камбия).
, 2016). Таким образом, любые отклонения функции TRW (N) от функции прямой пропорциональности (т.е., линейная функция с нулевым постоянным членом) были вызваны изменчивостью среднего D в годичном кольце. Эта изменчивость складывается из нескольких компонентов: (i) возрастных трендов ювенильной древесины (Высоцкая, Ваганов, 1989; Lei et al., 1996; Anfodillo et al., 2013), которые не учитываются в рассматриваемом наборе данных, (ii) унаследованный эффект, описываемый экспоненциальной функцией D ( N ), (iii) влияние текущих климатических колебаний во время расширения ячейки и (iv) стохастический компонент.Следовательно, функция TRW (N) имеет небольшую кривизну в области узких колец (рис. 3). Тем не менее, очень высокие коэффициенты детерминации, полученные для линейных моделей, свидетельствовали о высокой стабильности среднего размера клеток, образующихся у основания ствола дерева, на протяжении большей части жизни дерева. Напротив, близость этой зависимости утвердила TRW как параметр, интегрирующий влияние климатических колебаний преимущественно в период камбиальной деятельности и до ее начала (через состояние дерева в момент активизации камбия). Хотя погодные колебания влияют на анатомическое строение и функциональные особенности ксилемы, образующейся на следующих стадиях дифференцировки трахеид после окончания клеточного деления, их влияние практически не отразилось на TRW .
Хотя погодные колебания влияют на анатомическое строение и функциональные особенности ксилемы, образующейся на следующих стадиях дифференцировки трахеид после окончания клеточного деления, их влияние практически не отразилось на TRW .
Анатомические признаки обнаруживают меньшую вариабельность с ограниченным диапазоном значений по сравнению с клеточной продукцией, что в основном связано с функциональными ограничениями ксилемы как механической и водопроводной ткани (Fromm, 2013).Однако функциональная способность ксилемы чувствительна даже к небольшому изменению размеров клеток (например, способность проводить воду зависит от четвертой степени диаметра просвета; Tyree, Ewers, 1991). Такое функциональное ограничение, безусловно, может усложнить взаимосвязь анатомических признаков с другими признаками годичных колец и с климатическими переменными. Зависимость анатомических особенностей трахеид от их продукции прослеживается для всех деревьев и имеет общий и явно нелинейный характер. Эта нелинейность может быть выражена через значительную положительную связь, наблюдаемую между процессами продукции и последующей дифференцировки в случае небольшого числа клеток, где стрессовые условия во время клеточного деления трансформируются в образование лишь нескольких мелких и тонкостенных трахеид. Однако большая клеточная продукция ослабляет эту связь, и самые широкие кольца, образующиеся в благоприятных условиях, имеют анатомические черты, практически не зависящие от клеточной продукции.
Эта нелинейность может быть выражена через значительную положительную связь, наблюдаемую между процессами продукции и последующей дифференцировки в случае небольшого числа клеток, где стрессовые условия во время клеточного деления трансформируются в образование лишь нескольких мелких и тонкостенных трахеид. Однако большая клеточная продукция ослабляет эту связь, и самые широкие кольца, образующиеся в благоприятных условиях, имеют анатомические черты, практически не зависящие от клеточной продукции.
В широких кольцах как средние, так и максимальные оценки рассматриваемых параметров клеток колеблются вокруг значений, которые, вероятно, отражают оптимальную функциональность ксилемы для каждого отдельного дерева.Различия этих величин между деревьями выражают генетические или фенотипические (высота дерева, размерные характеристики кроны, корневая система) особенности конкретного дерева и микроусловия местообитания. Предположение о частичной генетической обусловленности строения ксилемы было подтверждено шкалами видов и происхождения несколькими исследованиями, например, количественным анализом анатомических признаков у сосен разного происхождения, имеющих общий ареал (Martín et al. , 2010; Matisons et al., 2019).Зависимость признаков КВА от вида наблюдалась в масштабе отдельных деревьев при сравнении структуры древесных колец привоя и подвоя в один и тот же год в гетеропрививках кедра сибирского на сосне обыкновенной (Дарикова и др., 2013). Очевидно, что генетические различия проявляются и в наименьшем масштабе отдельных генотипов. Размерные характеристики дерева, особенно его высота, доля тонких корней и фотосинтетический аппарат, определяют потребность растения в воде и приводят к ранее описанным тенденциям в анатомической структуре вдоль его вертикального и радиального уклонов, как сообщает Anfodillo. и другие.(2012, 2013) и других исследователей (Высоцкая, Ваганов, 1989; Lei et al., 1996; Mencuccini et al., 1997). Однако в данном исследовании влияние динамики высоты деревьев сведено к минимуму, на что указывает отсутствие многолетних трендов измеряемых характеристик клеток, а значит, имеет значение только разница в конечной высоте взрослых деревьев. Микроусловия определяют изменчивость микроклимата, наличие ресурсов для роста и развития деревьев и конкуренцию за ресурсы с соседями (Jaques, 1995; Van Do et al.
, 2010; Matisons et al., 2019).Зависимость признаков КВА от вида наблюдалась в масштабе отдельных деревьев при сравнении структуры древесных колец привоя и подвоя в один и тот же год в гетеропрививках кедра сибирского на сосне обыкновенной (Дарикова и др., 2013). Очевидно, что генетические различия проявляются и в наименьшем масштабе отдельных генотипов. Размерные характеристики дерева, особенно его высота, доля тонких корней и фотосинтетический аппарат, определяют потребность растения в воде и приводят к ранее описанным тенденциям в анатомической структуре вдоль его вертикального и радиального уклонов, как сообщает Anfodillo. и другие.(2012, 2013) и других исследователей (Высоцкая, Ваганов, 1989; Lei et al., 1996; Mencuccini et al., 1997). Однако в данном исследовании влияние динамики высоты деревьев сведено к минимуму, на что указывает отсутствие многолетних трендов измеряемых характеристик клеток, а значит, имеет значение только разница в конечной высоте взрослых деревьев. Микроусловия определяют изменчивость микроклимата, наличие ресурсов для роста и развития деревьев и конкуренцию за ресурсы с соседями (Jaques, 1995; Van Do et al. , 2015), которые также количественно влияют на оптимальную структуру древесины. Однако следует отметить, что индивидуальные различия не повлияли на принципиальный характер зависимостей D(N) и CWT (N). Все клетки зарождающейся ксилемы на момент их образования представляли размеры и первичные ЦВТ сходными с таковыми камбия, что дало возможность описать выявленные зависимости в виде экспоненциальных функций с числовыми коэффициентами явных значений.Экстремальные внешние условия подавляют скорость всех ростовых процессов (деления и дифференцировки клеток) в ксилеме одновременно. Предполагая общую нулевую точку для этих скоростей процессов роста, нулевое клеточное деление будет соответствовать нулевому расширению и нулевому отложению вторичной стенки. Следовательно, в точках пересечения функций с вертикальной осью (теоретический предел N → 0) трахеиды должны иметь те же размеры, что и исходные камбиальные клетки, не прошедшие процессы дифференцировки.Асимптотические значения D as и CWT as , к которым стремится кривая в области более широких колец ( N → max), характеризуют баланс между эффективностью и безопасностью оптимальной гидравлической архитектуры (размер просвета и прочность) для каждого дерева.
, 2015), которые также количественно влияют на оптимальную структуру древесины. Однако следует отметить, что индивидуальные различия не повлияли на принципиальный характер зависимостей D(N) и CWT (N). Все клетки зарождающейся ксилемы на момент их образования представляли размеры и первичные ЦВТ сходными с таковыми камбия, что дало возможность описать выявленные зависимости в виде экспоненциальных функций с числовыми коэффициентами явных значений.Экстремальные внешние условия подавляют скорость всех ростовых процессов (деления и дифференцировки клеток) в ксилеме одновременно. Предполагая общую нулевую точку для этих скоростей процессов роста, нулевое клеточное деление будет соответствовать нулевому расширению и нулевому отложению вторичной стенки. Следовательно, в точках пересечения функций с вертикальной осью (теоретический предел N → 0) трахеиды должны иметь те же размеры, что и исходные камбиальные клетки, не прошедшие процессы дифференцировки.Асимптотические значения D as и CWT as , к которым стремится кривая в области более широких колец ( N → max), характеризуют баланс между эффективностью и безопасностью оптимальной гидравлической архитектуры (размер просвета и прочность) для каждого дерева. Числовой коэффициент ( a ) регулирует диапазон существенной зависимости анатомических характеристик от клеточной продукции.
Числовой коэффициент ( a ) регулирует диапазон существенной зависимости анатомических характеристик от клеточной продукции.
Получены долгосрочные индексированные хронологии как независимые показатели воздействия климата на рост деревьев
Устранение устаревшего эффекта путем деления фактических измерений на модельные значения, процедура, аналогичная стандартизации хронологий ширины годичных колец, привела к получению индексированных хронологий для анатомических признаков.Значения этих индексированных хронологий были ближе к нормальному распределению по сравнению с первоначальными измерениями и, следовательно, показали большую пригодность для дендроклиматического анализа. Хотя представленные модели зависимостей от N демонстрируют признаки несовершенства, по крайней мере, для характеристик диаметра клеток, поскольку их индексированные хронологии имеют малозаметные долгосрочные тренды, отсутствующие в измеренных значениях. Необходимы дальнейшие исследования долгосрочной стабильности вариаций в анатомических хронологиях, чтобы улучшить этот простой подход к индексации, чтобы их можно было использовать в качестве многовековых заменителей для дендроклиматических реконструкций.
Индексированные хронологии были разработаны с меньшим количеством общего сигнала из-за устранения изменчивости, унаследованной от клеточного производства и внешних (климатических) процессов, записанных в нем. Однако существенные связи этих хронологий с климатическими факторами все же наблюдались. Независимость проиндексированных анатомических хронологий от процессов камбиальной активности нашла отражение в резких различиях в их реакции на климат по сравнению с климатическими сигналами, зарегистрированными в TRW , тесно связанном с N .В дальнейшем был получен набор стандартизированных хронологий, фиксирующих влияние климатических колебаний в течение сезона отдельно и независимо на все три этапа формирования ксилемы, т. е. (1) TRW как индикатор влияния условий внешней среды на клетки продукция в камбиальной зоне; 2) D max и D mean , объясняющие климатические условия в процессе расширения клеток в средней части зоны ранней древесины и во всем кольце соответственно; (3) CWT max и CWT mean , указывающие на климатическую изменчивость в процессе отложения вторичной клеточной стенки в типичных трахеидах поздней древесины и во всем кольце соответственно.
Сравнение откликов разработанных хронологий на климатические переменные выявило отчетливые различия в сезонности, интенсивности и даже направленности воздействия температуры и осадков на последовательные стадии развития древесины. Производство клеток продемонстрировало климатическую реакцию на доступность влаги, типичную для полузасушливых регионов, где положительная корреляция указывает на осадки как на источник воды, а отрицательная корреляция указывает на температуру как фактор высыхания (Liu et al., 2013; Бхандари и др., 2020). Однако, по крайней мере, для сосны изучаемой территории следует помнить, что крупные клетки ранней древесины составляют основную долю по количеству клеток. Это может быть связано как с видовой предрасположенностью, так и с типом климата изучаемой территории. В континентальном климате скорость камбиальной активности имеет одномодальную и асимметричную кинетику с максимумом ближе к началу сезона, совпадающему с началом развития ранней древесины (Попкова и др. , 2018; Fonti et al., 2020), что снижает долю поздней древесины сосны обыкновенной примерно до одной трети кольца, как это наблюдается у деревьев сосны обыкновенной, произрастающих в различных природных зонах региона (Жирнова и др., 2020, 2021). Следовательно, климатическая реакция TRW была больше сосредоточена на ранней древесине, чем на поздней древесине, а сезонный интервал значительных корреляций (т. е. с мая по первую половину июня) согласовывался с образованием трахеид ранней древесины. Различия в сезонности климатической реакции ранней и поздней древесины неоднократно отмечались разными исследователями при изучении ширины ранней и поздней древесины (De Grandpré et al., 2011; Кроуфорд и др., 2015 г.; Акоста-Эрнандес и др., 2019). Эти паттерны также применимы к продукции клеток в этих зонах по очевидным причинам.
, 2018; Fonti et al., 2020), что снижает долю поздней древесины сосны обыкновенной примерно до одной трети кольца, как это наблюдается у деревьев сосны обыкновенной, произрастающих в различных природных зонах региона (Жирнова и др., 2020, 2021). Следовательно, климатическая реакция TRW была больше сосредоточена на ранней древесине, чем на поздней древесине, а сезонный интервал значительных корреляций (т. е. с мая по первую половину июня) согласовывался с образованием трахеид ранней древесины. Различия в сезонности климатической реакции ранней и поздней древесины неоднократно отмечались разными исследователями при изучении ширины ранней и поздней древесины (De Grandpré et al., 2011; Кроуфорд и др., 2015 г.; Акоста-Эрнандес и др., 2019). Эти паттерны также применимы к продукции клеток в этих зонах по очевидным причинам.
Максимальный размер клеток в кольце зависит от количества осадков, выпадающих вскоре после начала вегетационного периода, когда доступность воды регулирует тургор и определяет скорость расширения, приводящую к образованию наиболее крупных трахеид, происходящих в это время (Passioura и Fry, 1992; Cabon et al. , 2020; типичное положение самой крупной клетки в середине ранней древесины см. на рис. 2).Также крупные трахеиды этой зоны сочетают интенсивные темпы и большую продолжительность экспансии клеток (Anfodillo et al., 2012; Cuny, 2013). Таким образом, даже если D max происходит из измерения только одной клетки, его значение в конкретном кольце может регистрировать наличие воды в течение нескольких недель. Межгодовые сдвиги дат, когда расширяется самая большая ячейка, обеспечивают дальнейшее увеличение внутрисезонного интервала, охватываемого климатическим сигналом в долговременной хронологии, примерно до длины месяца.Для среднего размера клетки аналогичная реакция на условия начала лета подготовила почву для отрицательных корреляций с обоими климатическими факторами в более позднее время сезона, что можно объяснить несколькими причинами. Например, отрицательная реакция на осадки может быть связана с большей долей мелких клеток поздней древесины в кольце, образующихся в более влажных условиях.
, 2020; типичное положение самой крупной клетки в середине ранней древесины см. на рис. 2).Также крупные трахеиды этой зоны сочетают интенсивные темпы и большую продолжительность экспансии клеток (Anfodillo et al., 2012; Cuny, 2013). Таким образом, даже если D max происходит из измерения только одной клетки, его значение в конкретном кольце может регистрировать наличие воды в течение нескольких недель. Межгодовые сдвиги дат, когда расширяется самая большая ячейка, обеспечивают дальнейшее увеличение внутрисезонного интервала, охватываемого климатическим сигналом в долговременной хронологии, примерно до длины месяца.Для среднего размера клетки аналогичная реакция на условия начала лета подготовила почву для отрицательных корреляций с обоими климатическими факторами в более позднее время сезона, что можно объяснить несколькими причинами. Например, отрицательная реакция на осадки может быть связана с большей долей мелких клеток поздней древесины в кольце, образующихся в более влажных условиях. Эта гипотеза была подтверждена явной временной стабильностью, наблюдаемой для начала этой конкретной реакции, которая была зафиксирована около летнего солнцестояния как триггерный фактор для формирования поздней древесины (ср.Drew and Downes, 2015, и ссылки там). Наоборот, повышенную температуру можно рассматривать как стимулятор развития растений за счет сокращения продолжительности отдельных стадий. Даже время наступления различных фенофаз для многих культурных растений рассчитано по суммам активных температур (Григорьева и др., 2010; Hatfield et al., 2011; Wypych et al., 2017). Поскольку продолжительность процесса расширения клеток больше влияет на окончательный радиальный размер, чем на его скорость (Anfodillo et al., 2012; Cuny, 2013), повышение температуры в сочетании с достаточной влажностью может приводить к уменьшению размеров клеток за счет более быстрого перехода клеток в стадию отложения вторичной стенки.
Эта гипотеза была подтверждена явной временной стабильностью, наблюдаемой для начала этой конкретной реакции, которая была зафиксирована около летнего солнцестояния как триггерный фактор для формирования поздней древесины (ср.Drew and Downes, 2015, и ссылки там). Наоборот, повышенную температуру можно рассматривать как стимулятор развития растений за счет сокращения продолжительности отдельных стадий. Даже время наступления различных фенофаз для многих культурных растений рассчитано по суммам активных температур (Григорьева и др., 2010; Hatfield et al., 2011; Wypych et al., 2017). Поскольку продолжительность процесса расширения клеток больше влияет на окончательный радиальный размер, чем на его скорость (Anfodillo et al., 2012; Cuny, 2013), повышение температуры в сочетании с достаточной влажностью может приводить к уменьшению размеров клеток за счет более быстрого перехода клеток в стадию отложения вторичной стенки.
Положительный отклик CWT на температуру и отрицательный отклик на осадки в течение мая наблюдался у CWT означает , но не у CWT max , что можно интерпретировать как раннюю древесину, являющуюся регистратором этого сигнала. . Формирование более мелких и толстых клеток ранней древесины в сухих и жарких условиях может быть частью механизма адаптации растениями структуры ранней древесины к засухе, обеспечивая более высокую безопасность водообеспечения (снижение риска кавитации и эмболии) через перераспределение доступные ресурсы от расширения клеток до осаждения стенок.Это явление в период наиболее сильных засух приводит к формированию ИАДФ в виде полосы позднедеревоподобных трахеид в ранней древесине (Lautner, 2013; Battipaglia et al., 2016; De Micco et al., 2016). Однако требование достаточной механической прочности стенок трахеид более мелких клеток поздней древесины обычно выполнялось без наличия такой реакции.
. Формирование более мелких и толстых клеток ранней древесины в сухих и жарких условиях может быть частью механизма адаптации растениями структуры ранней древесины к засухе, обеспечивая более высокую безопасность водообеспечения (снижение риска кавитации и эмболии) через перераспределение доступные ресурсы от расширения клеток до осаждения стенок.Это явление в период наиболее сильных засух приводит к формированию ИАДФ в виде полосы позднедеревоподобных трахеид в ранней древесине (Lautner, 2013; Battipaglia et al., 2016; De Micco et al., 2016). Однако требование достаточной механической прочности стенок трахеид более мелких клеток поздней древесины обычно выполнялось без наличия такой реакции.
В отличие от сосновых насаждений, произрастающих в центре долины, полностью в пределах засушливой степной зоны с возможной генетической адаптацией к частым засухам, выборочная популяция горного леса в данном исследовании могла быть адаптирована к любой конкретной климатической экстремальности только за счет фенотипической пластичности, из-за смешения генофонда между деревьями, растущими на различных высотах и склонах (ср. Олден и Лупстра, 1987 г .; Ларионова и др., 2007; Чанг и др., 2020). Поэтому, как и в других горных местообитаниях региона (см. Белокопытова и др., 2020), CWT в поздней древесине выражали положительную реакцию на осадки и отрицательную обратную связь с температурой к концу сезона, что отражалось как на CWT max и CWT означают хронологии.
Олден и Лупстра, 1987 г .; Ларионова и др., 2007; Чанг и др., 2020). Поэтому, как и в других горных местообитаниях региона (см. Белокопытова и др., 2020), CWT в поздней древесине выражали положительную реакцию на осадки и отрицательную обратную связь с температурой к концу сезона, что отражалось как на CWT max и CWT означают хронологии.
Региональное потепление в исследуемом регионе, очевидно, увеличило продолжительность сезона с температурами, подходящими для образования древесины, как весной, так и осенью (см. Rogers, Mosely-Thompson, 1995; Росгидромет, 2014).Очень показательно, что индексированные анатомические хронологии гораздо более четко и отчетливо отреагировали на этот сдвиг, чем TRW хронология, как показал анализ временной стабильности климатических реакций. Хронологии D указывали на явно более раннее начало ксилогенеза, тогда как конечная точка значимых дендроклиматических корреляций для CWT сместилась на более поздние календарные даты. В то же время закономерности реакции в середине сезона, отражающие переход от ранней древесины к поздней, оставались «приклеенными» к одной и той же календарной дате во второй половине июня, что доказывает, что длина дня является основной движущей силой этого перехода для Сосна обыкновенная.В целом стабильная интенсивность и равномерные долговременные сдвиги климатического сигнала в индексированных анатомических хронологиях поддерживают их использование в качестве мелкомасштабных климатических прокси, несмотря на снижение общего сигнала при индексации.
В то же время закономерности реакции в середине сезона, отражающие переход от ранней древесины к поздней, оставались «приклеенными» к одной и той же календарной дате во второй половине июня, что доказывает, что длина дня является основной движущей силой этого перехода для Сосна обыкновенная.В целом стабильная интенсивность и равномерные долговременные сдвиги климатического сигнала в индексированных анатомических хронологиях поддерживают их использование в качестве мелкомасштабных климатических прокси, несмотря на снижение общего сигнала при индексации.
Выводы
В этом исследовании, в то время как ширина годичных колец демонстрировала линейность, почти прямо пропорциональную количеству клеток, клеточные признаки расшифровывали нелинейность гидравлических и механических функций ксилемы в отношении продукции клеток и демонстрировали более сильную связь в областях узких колец и стабильная анатомическая структура более широких колец.Эти отношения описывались отрицательной экспоненциальной функцией. Кроме того, взаимосвязь между климатом и ростом показала, что на стандартизированную индексированную хронологию признаков древесины сильно влияют климатические факторы, особенно наличие влаги. Это отражает применимость данного исследования роста растений и качества древесины в сценариях изменения климата, особенно в условиях засухи. Однако конкретные механизмы и причинно-следственный характер наблюдаемых климатических реакций необходимо дополнительно изучить с точки зрения физиологии деревьев этого и других видов, чтобы получить более надежную базу данных для проведения убедительных наблюдений.Временная стабильность выявленных взаимосвязей также должна быть исследована в масштабе столетия, поскольку длина представленного набора данных позволяет такое исследование. Независимо от направления будущих исследований, полученные в этом исследовании анатомические хронологии, чувствительные к климату, предлагают важную черту четкого временного внутрисезонного разделения климатического сигнала. Увеличение периода охвата хронологий используемых наборов анатомических данных делает их перспективными прокси для обратной дендроклиматической задачи, что может привести к реконструкции важных климатических факторов для относительно коротких внутрисезонных интервалов.
Кроме того, взаимосвязь между климатом и ростом показала, что на стандартизированную индексированную хронологию признаков древесины сильно влияют климатические факторы, особенно наличие влаги. Это отражает применимость данного исследования роста растений и качества древесины в сценариях изменения климата, особенно в условиях засухи. Однако конкретные механизмы и причинно-следственный характер наблюдаемых климатических реакций необходимо дополнительно изучить с точки зрения физиологии деревьев этого и других видов, чтобы получить более надежную базу данных для проведения убедительных наблюдений.Временная стабильность выявленных взаимосвязей также должна быть исследована в масштабе столетия, поскольку длина представленного набора данных позволяет такое исследование. Независимо от направления будущих исследований, полученные в этом исследовании анатомические хронологии, чувствительные к климату, предлагают важную черту четкого временного внутрисезонного разделения климатического сигнала. Увеличение периода охвата хронологий используемых наборов анатомических данных делает их перспективными прокси для обратной дендроклиматической задачи, что может привести к реконструкции важных климатических факторов для относительно коротких внутрисезонных интервалов.
Заявление о доступности данных
Оригинальные материалы, представленные в исследовании, включены в статью/дополнительный материал, дальнейшие запросы можно направлять соответствующему автору.
Вклад авторов
EV задумал и разработал исследование. EB контролировал и координировал исследование в целом. EV и EB оказали финансовую поддержку. Э.В., Э.Б., Д.З. и Л.Б. проводили полевые работы. ДЗ руководил и участвовал в обработке образцов, руководил анатомическими измерениями.ДД и МЗ под руководством ЛБ выполняли расчеты, анализ данных и создавали графики. LB, KU и ST завершили рукопись, а затем отредактировали ее. Все авторы внесли свой вклад в интерпретацию результатов и написание первого проекта.
Финансирование
Работа выполнена при финансовой поддержке Министерства науки и высшего образования Российской Федерации, код научной темы ФСРЗ-2020-0010, и РФФИ, грант № 20-016-00049. Источники финансирования не участвовали в разработке дизайна исследования, в сборе, анализе, интерпретации данных, в написании и в решении представить статью для публикации.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все претензии, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Авторы выражают благодарность техническим сотрудникам Лаборатории дендроэкологии и экологического мониторинга (Хакасский технический институт, СФУ, Россия) за кропотливую работу по обработке образцов и анатомическим измерениям. Также ценят советы проф. В.В. Шишова (Сибирский федеральный университет, Россия) в области статистического анализа.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www. frontiersin.org/articles/10.3389/fpls.2021.719796/full#supplementary-material
frontiersin.org/articles/10.3389/fpls.2021.719796/full#supplementary-material
Каталожные номера
Акоста-Эрнандес, А.С., Камареро, Дж.Дж., и Помпа-Гарсия, М. (2019). Реакция сезонного роста на климат во влажных и сухих хвойных лесах. ИАВА J . 40, 311–С1. дои: 10.1163/22941932-401
Полнотекстовая перекрестная ссылка | Академия Google
Олден, Дж., и Лупстра, К. (1987). Генетическое разнообразие и структура популяции Picea glauca на высотном градиенте во внутренних районах Аляски. Кан. Дж. Для. Рез . 17, 1519–1526. дои: 10.1139/x87-234
Полнотекстовая перекрестная ссылка | Академия Google
Алисов Б.П. (1956). Климат СССР . Москва: МГУ.
Алони, Р. (2021). Сосудистая дифференциация и растительные гормоны . Чам: Спрингер.
Анфодильо Т., Делорье А., Менарди Р., Тедольди Л., Пети Г. и Росси С. (2012). Расширение каналов ксилемы в хвойном дереве зависит от более длительного времени распространения клеток вниз по стволу. Дж. Экспл. Бот . 63, 837–845. дои: 10.1093/jxb/err309
Дж. Экспл. Бот . 63, 837–845. дои: 10.1093/jxb/err309
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Анфодильо Т., Пети Г. и Кривелларо А. (2013). «Расширение осевого канала у древесных пород: до сих пор игнорируемая анатомическая модель», в «Структура древесины в биологии и экологии растений», , изд. П. Баас (Лейден: Брилл), 24–36.
Академия Google
Арзак, А., Лопес-Сеперо, Х.М., Бабушкина, Е.А., и Гомес, С. (2018). Применение методов подготовки твердых тканей к анатомии древесины: Визуализация полированных образцов, залитых полиметилметакрилатом. Дендрохронология 51, 76–81. doi: 10.1016/j.dendro.2018.08.005
Полнотекстовая перекрестная ссылка | Академия Google
Battipaglia, G., Campelo, F., Vieira, J., Grabner, M., De Micco, V., Nabais, C., et al. (2016). Структура и функция внутригодовых колебаний плотности: обратите внимание на пробелы. Фронт. Завод Наука . 7:595. doi: 10. 3389/fpls.2016.00595
3389/fpls.2016.00595
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Белокопытова Л.В., Фонти П., Бабушкина Е.А., Жирнова Д.Ф., Ваганов Е.А. (2020). Свидетельства различной засухоустойчивости процессов развития клеток ксилемы южно-сибирской сосны. Леса 11:294. дои: 10.3390/f11121294
Полнотекстовая перекрестная ссылка | Академия Google
Бхандари, С., Спир, Дж. Х., Хан, А., и Ахмед, М. (2020). Сигнал засухи в годичных кольцах трех видов хвойных деревьев из Северного Пакистана. Дендрохронология 63:125742. doi: 10.1016/j.dendro.2020.125742
Полнотекстовая перекрестная ссылка | Академия Google
Кабон, А., Фернандес-де-Унья, Л., Хеа-Искьердо, Г., Мейнцер, Ф. К., Вудрафф, Д. Р., Мартинес-Вилальта, Дж., и др. (2020). Контроль водного потенциала вызванного тургором увеличения трахеид у сосны обыкновенной на краю ее ксерического распространения. Новый Фитол . 225, 209–221. doi: 10.1111/nph. 16146
16146
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Каррер М., Кастаньери Д., Прендин А. Л., Пети Г.и фон Аркс, Г. (2017). Ретроспективный анализ анатомических признаков древесины показывает недавнее расширение камбиальной активности деревьев у двух высокогорных хвойных. Фронт. Завод Наука . 8:737. doi: 10.3389/fpls.2017.00737
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кастаньери Д., Баттипалья Г., фон Аркс Г., Пачеко А. и Каррер М. (2018). Анатомия годичных колец и соотношение изотопов углерода показывают как прямое, так и унаследованное влияние климата на формирование бимодальной ксилемы у Pinus pinea . Физиол дерева . 38, 1098–1109. doi: 10.1093/treephys/tpy036
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кастаньери Д., Фонти П., фон Аркс Г. и Каррер М. (2017). Как климат влияет на морфогенез ксилемы в течение вегетационного периода? выводы из долгосрочной анатомии внутреннего кольца Picea abies. Энн. Бот . 119, 1011–1020. doi: 10.1093/aob/mcw274
Энн. Бот . 119, 1011–1020. doi: 10.1093/aob/mcw274
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чанг, М.Ю., Сон С., Эррандо-Морайра С., Танг С.К., Маки М., Ким Ю.Д. и др. (2020). Включение различий между генетическим разнообразием деревьев и травянистых растений в стратегии сохранения. Консерв. Биол . 34, 1142–1151. doi: 10.1111/cobi.13467
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кук, Э. Р., и Кайрюкстис, Л. А. (1990). Методы дендрохронологии. Применение в науках об окружающей среде. Дордрехт: Спрингер.
Академия Google
Кук, Э.Р. и Питерс К. (1981). Сглаживающий сплайн: новый подход к стандартизации рядов ширины внутренних годичных колец леса для дендроклиматических исследований. Кольцевой бык . 41, 45–53.
Академия Google
Кроуфорд, С. Дж., Гриффин, Д., и Кипфмюллер, К. Ф. (2015). Регистрация сезонных сигналов осадков в северной части Скалистых гор, США, с использованием годичных колец ранней и поздней древесины. Ж. Геофиз. Рез. Биогеоски . 120, 428–440. дои: 10.1002/2014JG002740
Ж. Геофиз. Рез. Биогеоски . 120, 428–440. дои: 10.1002/2014JG002740
Полнотекстовая перекрестная ссылка | Академия Google
Куни, Х.Э. (2013). Внутригодовая динамика формирования древесины трех пород хвойных (пихты европейской, ели европейской и сосны обыкновенной) на северо-востоке Франции: от описания сезонных закономерностей роста к изучению влияния среды на кинетику развития клеток и анатомический Особенности Xylem . [Диссертация]. Нэнси: Университет Лотарингии (на французском языке).
Cuny, H.E., Fonti, P., Rathgeber, C.B., von Arx, G., Peters, R.L., and Frank, D.C. (2019). Связи в кинетике дифференцировки клеток смягчают влияние температуры воздуха на анатомию древесины хвойных пород. Окружающая среда клеток растений . 42, 1222–1232. doi: 10.1111/pce.13464
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cuny, HE, and Rathgeber, CB (2016). Ксилогенез: хвойные деревья умеренных лесов слушают климатическую сказку в течение вегетационного периода, но помнят только последние слова! Физика растений . 171, 306–317. doi: 10.1104/pp.16.00037
171, 306–317. doi: 10.1104/pp.16.00037
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Куни, Х.Э., Ратгебер, К.Б., Франк, Д., Фонти, П., и Фурнье, М. (2014). Кинетика развития трахеид объясняет структуру годичных колец хвойных деревьев. Новый Фитол . 203, 1231–1241. doi: 10.1111/nph.12871
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дарикова Ю.А., Ваганов Е.А., Кузнецова Г.В., Грачев А.М. (2013). Изменения анатомического строения годичных колец подвоя и привоя гетеропривоев кедра сибирского. Деревья 27, 1621–1631.doi: 10.1007/s00468-013-0909-6
Полнотекстовая перекрестная ссылка | Академия Google
De Grandpré, L., Tardif, J.C., Hessl, A., Pederson, N., Conciatori, F., Green, T.R., et al. (2011). Сезонный сдвиг в климатических реакциях деревьев Pinus sibirica , Pinus sylvestris и Larix sibirica из полузасушливых районов северо-центральной Монголии. Кан. Дж. Для. Рез . 41, 1242–1255. дои: 10.1139/x11-051
Кан. Дж. Для. Рез . 41, 1242–1255. дои: 10.1139/x11-051
Полнотекстовая перекрестная ссылка | Академия Google
Де Микко, В., Campelo, F., De Luis, M., Bräuning, A., Grabner, M., Battipaglia, G., et al. (2016). Внутригодовые колебания плотности годичных колец: как, когда, где и почему? ИАВА J . 37, 232–259. дои: 10.1163/22941932-20160132
Полнотекстовая перекрестная ссылка | Академия Google
De Micco, V., Carrer, M., Rathgeber, C.B., Camarero, J.J., Voltas, J., Cherubini, P., et al. (2019). От ксилогенеза до годичных колец: признаки древесины для изучения реакции деревьев на изменения окружающей среды. ИАВА J .40, 155–182. дои: 10.1163/22941932-401
Полнотекстовая перекрестная ссылка | Академия Google
Дуглас, А.Э. (1925). Годичные кольца и климат. науч. Пн . 21, 95–99.
Академия Google
Дрю, Д.М., и Даунс, Г.М. (2015). Модель роста ствола и формирования древесины у Pinus radiata . Деревья 29, 1395–1413. doi: 10.1007/s00468-015-1216-1
Деревья 29, 1395–1413. doi: 10.1007/s00468-015-1216-1
Полнотекстовая перекрестная ссылка | Академия Google
Эспер Дж., Крушич П.J., Ljungqvist, F.C., Luterbacher, J., Carrer, M., Cook, E., et al. (2016). Рейтинг температурных реконструкций прошлого тысячелетия на основе годичных колец. Кв. науч. Версия . 145, 134–151. doi: 10.1016/j.quascirev.2016.05.009
Полнотекстовая перекрестная ссылка | Академия Google
Фонти, М. В., Бабушкина, Е. А., Жирнова, Д. Ф., и Ваганов, Е. А. (2020). Ксилогенез сосны обыкновенной в разновозрастном насаждении Минусинской котловины (Южная Сибирь). Ж. Сиб. Кормили. ун-тБиол . 13, 197–207. дои: 10.17516/1997-1389-0323
Полнотекстовая перекрестная ссылка | Академия Google
Фриттс, ХК (1976). Годичные кольца деревьев и климат . Лондон: Академическая пресса.
Академия Google
Фромм, Дж. (2013). Клеточные аспекты формирования древесины. Берлин, Гейдельберг: Springer.
Академия Google
Фрай Г. и Чок Л. (1957). Изменение плотности древесины Pinus patula , выращенного в Кении. Лесное хозяйство 30, 29–45. doi: 10.1093/oxfordjournals.forestry.a063093
Полнотекстовая перекрестная ссылка | Академия Google
Гебрегеоргис, Э. Г., Бонецка, Дж., Пятковски, М., Робертсон, И., и Ратгебер, К. (2021). SabaTracheid 1.0: новая программа для количественного анализа анатомии древесины хвойных пород — демонстрация африканского можжевельника в бассейне Голубого Нила. Фронт. Завод Наука . 12:21. doi: 10.3389/fpls.2021.595258
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Григорьева Е.А., Мацаракис, А., и Де Фрейтас, Ч.Р. (2010). Анализ прироста градусо-дней как индикатора воздействия на климат в регионе с экстремальной годовой амплитудой температуры воздуха. Клим. Рез . 42, 143–154. дои: 10.3354/cr00888
Полнотекстовая перекрестная ссылка | Академия Google
Hatfield, J. L., Boote, K.J., Kimball, B.A., Ziska, L.H., Izaurralde, R.C., Ort, D., et al. (2011). Воздействие климата на сельское хозяйство: последствия для растениеводства. Агрон. Дж . 103, 351–370.doi: 10.2134/agronj2010.0303
L., Boote, K.J., Kimball, B.A., Ziska, L.H., Izaurralde, R.C., Ort, D., et al. (2011). Воздействие климата на сельское хозяйство: последствия для растениеводства. Агрон. Дж . 103, 351–370.doi: 10.2134/agronj2010.0303
Полнотекстовая перекрестная ссылка | Академия Google
Холмс, Р.Л. (1983). Компьютерный контроль качества при датировании и измерении годичных колец. Кольцевой бык . 43, 69–78.
Академия Google
Хьюз, М.К., Светнам, Т.В., и Диаз, Х.Ф. (2010). Дендроклиматология: достижения и перспективы . Дордрехт: Спрингер.
Академия Google
Жак, К. Э. (1995). Взаимосвязь между формой ствола, сомкнутостью насаждения и условиями участка: влияние условий окружающей среды на аллометрию деревьев и структуру леса в западно-центральной части Альберты .[Диссертация магистра наук]. Сент-Катаринс: Университет Брока.
Академия Google
Каптейн, Дж. К. (1914). Рост деревьев и метеорологические факторы. Recueil des Travaux Botaniques Néerlandais 11, 70–93.
Recueil des Travaux Botaniques Néerlandais 11, 70–93.
Академия Google
Кирдянов А., Хьюз М., Ваганов Э., Швайнгрубер Ф. и Силкин П. (2003). Значение температуры начала лета и даты схода снега для роста деревьев в сибирской субарктике. Деревья 17, 61–69. дои: 10.1007/с00468-002-0209-з
Полнотекстовая перекрестная ссылка | Академия Google
Ларионова А.Ю., Экарт А.К. и Кравченко А.Н. (2007). Генетическое разнообразие и структура популяций пихты сибирской ( Abies sibirica Ledeb .) в Средней Сибири, Россия. Евразийский J. For. Рез . 10, 185–192.
Академия Google
Ларсон, П.Р. (1994). Сосудистый камбий. Развитие и структура. Берлин, Гейдельберг: Springer.
Академия Google
Лотнер, С.(2013). «Формирование древесины в условиях засухи и засоления», в Cellular Aspects of Wood Formation , ed. Дж. Фромм. (Берлин: Springer), 187–202.
Академия Google
Лей, Х. , Милота, М. Р., и Гартнер, Б. Л. (1996). Различия между и внутри деревьев в анатомии и удельном весе древесины дуба белого Орегона ( Quercus garryana Dougl.). ИАВА J . 17, 445–461. дои: 10.1163/22941932-
, Милота, М. Р., и Гартнер, Б. Л. (1996). Различия между и внутри деревьев в анатомии и удельном весе древесины дуба белого Орегона ( Quercus garryana Dougl.). ИАВА J . 17, 445–461. дои: 10.1163/22941932-
642
Полнотекстовая перекрестная ссылка | Академия Google
Лю, Х., Park Williams, A., Allen, C.D., Guo, D., Wu, X., Anenkhonov, O.A., et al. (2013). Быстрое потепление ускоряет снижение роста деревьев в полузасушливых лесах Внутренней Азии. Глоб. Сменить Биол . 19, 2500–2510. doi: 10.1111/gcb.12217
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ljungqvist, F.C., Piermattei, A., Seim, A., Krusic, P.J., Büntgen, U., He, M., et al. (2020). Рейтинг реконструкций гидроклимата прошлого тысячелетия на основе годичных колец. Кв.науч. Версия . 230:106074. doi: 10.1016/j.quascirev.2019.106074
Полнотекстовая перекрестная ссылка | Академия Google
Мартин, Дж. А., Эстебан, Л. Г., Де Паласиос, П. , и Фернандес, Ф. Г. (2010). Различия в анатомических признаках древесины Pinus sylvestris L. между испанскими регионами происхождения. Деревья 24, 1017–1028. doi: 10.1007/s00468-010-0471-4
, и Фернандес, Ф. Г. (2010). Различия в анатомических признаках древесины Pinus sylvestris L. между испанскими регионами происхождения. Деревья 24, 1017–1028. doi: 10.1007/s00468-010-0471-4
Полнотекстовая перекрестная ссылка | Академия Google
Мартин-Бенито Д., Анчукайтис К.Дж., Эванс М.Н., дель Рио М., Бекман Х. и Канельяс И. (2017). Влияние засухи на анатомию ксилемы и эффективность водопользования двух сосуществующих видов сосны. Леса 8:332. дои: 10.3390/f80
Полнотекстовая перекрестная ссылка | Академия Google
Матисонс Р., Кришанс О., Карклина А., Адамович А., Янсонс А. и Гартнер Х. (2019). Пластичность и климатическая чувствительность анатомии древесины способствуют продуктивности восточно-балтийских происхождений сосны обыкновенной. Для. Экол.Управление . 452:117568. doi: 10.1016/j.foreco.2019.117568
Полнотекстовая перекрестная ссылка | Академия Google
Менкучини, М., Грейс, Дж. , и Фиораванти, М. (1997). Биомеханические и гидравлические детерминанты строения дерева сосны обыкновенной: анатомическая характеристика. Физиол дерева . 17, 105–113. doi: 10.1093/treephys/17.2.105
, и Фиораванти, М. (1997). Биомеханические и гидравлические детерминанты строения дерева сосны обыкновенной: анатомическая характеристика. Физиол дерева . 17, 105–113. doi: 10.1093/treephys/17.2.105
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Олано, Дж. М., Эудженио, М., Гарсия-Сервигон, А. И., Фолч, М.и Розас, В. (2012). Количественная анатомия трахеид выявляет сложный экологический контроль структуры древесины в континентальном средиземноморском климате. Междунар. J. Plant Sci . 173, 137–149. дои: 10.1086/663165
Полнотекстовая перекрестная ссылка | Академия Google
Панюшкина П., Хьюз М., Ваганов Э. и Манро М. (2003). Летняя температура на северо-востоке Сибири с 1642 г., реконструированная по размерам трахеид и количеству клеток Larix cajanderi . Кан.Дж. Для. Рез . 33, 1905–1914. дои: 10.1139/x03-109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пассиура, Дж. Б., и Фрай, С. К. (1992). Тургор и расширение клеток: за пределами уравнения Локхарта. австр. Дж. Завод Физиол . 19, 565–576. дои: 10.1071/PP9920565
Б., и Фрай, С. К. (1992). Тургор и расширение клеток: за пределами уравнения Локхарта. австр. Дж. Завод Физиол . 19, 565–576. дои: 10.1071/PP9920565
Полнотекстовая перекрестная ссылка | Академия Google
Peters, R.L., Balanzategui, D., Hurley, A.G., von Arx, G., Prendin, A.L., Cuny, H.E., et al. (2018). RAPTOR: Органайзер трахеид ряда и положения в R. Дендрохронология 47, 10–16. doi: 10.1016/j.dendro.2017.10.003
Полнотекстовая перекрестная ссылка | Академия Google
Piermattei, A., von Arx, G., Avanzi, C., Fonti, P., Gärtner, H., Piotti, A., et al. (2020). Функциональные связи анатомических признаков древесины ели европейской. Фронт. Завод Наука . 11:683. doi: 10.3389/fpls.2020.00683
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пломион К., Лепровост Г. и Стоукс А.(2001). Образование древесины у деревьев. Завод Физиол . 127, 1513–1523. doi: 10.1104/стр.010816
Полнотекстовая перекрестная ссылка | Академия Google
Попкова М. , Ваганов Е. А., Шишов В. В., Бабушкина Е. А., Росси С., Брюханова М. В. и др. (2018). Смоделированные трахеидограммы показывают влияние засухи на структуру годичных колец Pinus sylvestris из сибирской лесостепи. Фронт. Завод Наука . 9:1144. doi: 10.3389/fpls.2018.01144
, Ваганов Е. А., Шишов В. В., Бабушкина Е. А., Росси С., Брюханова М. В. и др. (2018). Смоделированные трахеидограммы показывают влияние засухи на структуру годичных колец Pinus sylvestris из сибирской лесостепи. Фронт. Завод Наука . 9:1144. doi: 10.3389/fpls.2018.01144
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Прицков, К., Генрих И., Грудд Х. и Хелле Г. (2014). Взаимосвязь между анатомией древесины, шириной годичных колец и плотностью древесины Pinus sylvestris L. и климатом в высоких широтах на севере Швеции. Дендрохронология 32, 295–302. doi: 10.1016/j.dendro.2014.07.003
Полнотекстовая перекрестная ссылка | Академия Google
Ринн, Ф. (2003). TSAP-Win: Анализ и представление временных рядов для дендрохронологии и связанных приложений: Справочник пользователя . Гейдельберг: РИННТЕХ.
Академия Google
Роджерс, Дж. К., и Мозли-Томпсон, Э. (1995). Атлантические арктические циклоны и мягкие сибирские зимы 1980-х гг. Геофиз. Рез. Письмо . 22, 799–802. дои: 10.1029/95GL00301
Геофиз. Рез. Письмо . 22, 799–802. дои: 10.1029/95GL00301
Полнотекстовая перекрестная ссылка
Росгидромета (2014). Второй оценочный отчет Росгидромета об изменении климата и его последствиях в Российской Федерации . Москва, Росгидромет.
Академия Google
Швайнгрубер, Ф. Х. (2012). Годичные кольца: основы и приложения дендрохронологии . Дордрехт: Спрингер.
Академия Google
Сео, Дж. В., Смильянич, М., и Уилмкинг, М. (2014). Оптимизация клеточно-анатомических хронологий сосны обыкновенной путем пошагового увеличения количества включенных радиальных рядов трахеид — тематическое исследование на основе трех скандинавских памятников. Дендрохронология 32, 205–209. doi: 10.1016/j.dendro.2014.02.002
Полнотекстовая перекрестная ссылка | Академия Google
Силкин П.П. (2010). Методы многопараметрического анализа структуры годичных колец хвойных пород . Красноярск: Сибирский федеральный университет.
Соуто-Эрреро, М., Розас, В., и Гарсия-Гонсалес, И. (2017). 481-летняя хронология дубовых сосудов ранней древесины как независимый от возраста климатический показатель в северо-западной Иберии. Глоб. Планета. Изменение 155, 20–28. doi: 10.1016/j.gloplacha.2017.06.003
Полнотекстовая перекрестная ссылка | Академия Google
Стэнглер, Д. Ф., Кале, Х.П., Раден М., Ларыш Э., Зайферт Т. и Спикер Х. (2021). Влияние внутрисезонной засухи на кинетику дифференциации трахеид и динамику сезонного роста ели европейской на градиенте высот. Леса 12:274. дои: 10.3390/f12030274
Полнотекстовая перекрестная ссылка | Академия Google
Тайри, М.Т., и Эверс, Ф.В. (1991). Гидравлическая архитектура деревьев и других древесных растений. Новый Фитол . 119, 345–360. doi: 10.1111/j.1469-8137.1991.tb00035.x
Полнотекстовая перекрестная ссылка | Академия Google
Упадхьяй К.К., Шах С.К., Рой А. и Трипати С.К. (2021). Дендроклиматология тика указывает на преобладающие климатические условия тропических влажных лесов Индии. Экол. Индикация . 129:107888. doi: 10.1016/j.ecolind.2021.107888
Экол. Индикация . 129:107888. doi: 10.1016/j.ecolind.2021.107888
Полнотекстовая перекрестная ссылка | Академия Google
Ваганов Е.А. (1990). «Метод трахеидограммы в анализе годичных колец и его применение», в Методах дендрохронологии.Применение в науках об окружающей среде , ред. Э. Р. Кук, Л. А. Кайрюкстис. (Дордрехт: Kluwer Acad. Publ.), 63–75.
Академия Google
Ваганов, Э. А., Анчукайтис, К. Дж., и Эванс, М. (2011). «Насколько хорошо изучены процессы, создающие дендроклиматические записи? механистическая модель климатического контроля динамики годичных колец хвойных деревьев», в Дендроклиматология: прогресс и перспективы , под ред. М.К. Хьюз, Т.В. Светнам, Х. Ф. Диас (Дордрехт: Springer), 37–75.
Академия Google
Ваганов Е.А., Бабушкина Е.А., Белокопытова Л.В., Жирнова Д.Ф. (2020). Небольшие колебания толщины клеточных стенок в ксилеме сосны и ели: сигнал от камбия? PLoS ONE 15: E 0233106. doi: 10.1371/journal.pone.0233106
doi: 10.1371/journal.pone.0233106
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ваганов Э.А., Хьюз М.К., Шашкин А.В. (2006). Динамика роста годичных колец хвойных деревьев: образы среды прошлого и будущего .Дордрехт: Спрингер.
Академия Google
Ваганов Е.А., Шашкин А.В., Свидерская Л.В., Высоцкая Л.Г. (1985). Гистометрический анализ роста древесных растений . Новосибирск: Наука.
Ван До Т., Сато Т., Сайто С., Козан О., Ямагава Х., Нагамацу Д. и др. (2015). Влияние микротопографии на структуру древостоя и видовое разнообразие деревьев в старовозрастном вечнозеленом широколиственном лесу на юго-западе Японии. Глоб.Экол. Сохраняйте . 4, 185–196. doi: 10.1016/j.gecco.2015.06.010
Полнотекстовая перекрестная ссылка | Академия Google
Виейра, Дж., Карвалью, А., и Кампелу, Ф. (2020). Рост деревьев при изменении климата: данные о сроках и кинетике ксилогенеза. Фронт. Завод Наука . 11:90. doi: 10.3389/fpls.2020.00090
11:90. doi: 10.3389/fpls.2020.00090
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
фон Аркс, Г., и Каррер, М. (2014). ROXAS — новый инструмент для построения многовековых хронологий просвета трахеид у хвойных. Дендрохронология 32, 290–293. doi: 10.1016/j.dendro.2013.12.001
Полнотекстовая перекрестная ссылка | Академия Google
фон Аркс Г., Кривелларо А., Прендин А. Л., Куфар К. и Каррер М. (2016). Количественная анатомия древесины – практические рекомендации. Фронт. Завод Наука . 7:781. doi: 10.3389/fpls.2016.00781
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Высоцкая Л.Г., Ваганов Е.А. (1989). Компоненты изменчивости радиальных размеров клеток в годичных кольцах хвойных. ИАВА J . 10, 417–426. дои: 10.1163/22941932-134
Полнотекстовая перекрестная ссылка | Академия Google
Weiskopf, S.R., Rubenstein, M.A., Crozier, L.G., Gaichas, S. , Griffis, R., Halofsky, J.E., et al. (2020). Влияние изменения климата на биоразнообразие, экосистемы, экосистемные услуги и управление природными ресурсами в Соединенных Штатах. науч. Общая окружающая среда . 733:137782. doi: 10.1016/j.scitotenv.2020.137782
, Griffis, R., Halofsky, J.E., et al. (2020). Влияние изменения климата на биоразнообразие, экосистемы, экосистемные услуги и управление природными ресурсами в Соединенных Штатах. науч. Общая окружающая среда . 733:137782. doi: 10.1016/j.scitotenv.2020.137782
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Выпич А., Суликовска А., Устрнул З. и Чекерда Д. (2017). Изменчивость роста градусо-дней в Польше в ответ на продолжающиеся климатические изменения в Европе. Интерн. Дж. Биометеорол . 61, 49–59. doi: 10.1007/s00484-016-1190-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сюй С., Лю Х., Аненхонов О. А., Королюк А. Ю., Санданов Д. В., Балсанова Л. Д. и соавт. (2017). Долгосрочная устойчивость лесов к изменению климата проявляется гибелью, возобновлением и ростом в полузасушливой южной Сибири. Глоб. Сменить Биол . 23, 2370–2382. doi: 10.1111/gcb.13582
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ясуэ К. , Фунада Р., Кобаяши О. и Отани Дж. (2000). Влияние размеров трахеид на вариации максимальной плотности Picea glehnii и связь с климатическими факторами. Деревья 14, 223–229. дои: 10.1007/PL00009766
, Фунада Р., Кобаяши О. и Отани Дж. (2000). Влияние размеров трахеид на вариации максимальной плотности Picea glehnii и связь с климатическими факторами. Деревья 14, 223–229. дои: 10.1007/PL00009766
Полнотекстовая перекрестная ссылка | Академия Google
Цахнер, Р., Лотан, Дж.E. и Baughman, WD (1964). Особенности ранней и поздней древесины красной сосны, выращенной в условиях имитации засухи и орошения. Для. наука . 10, 361–370. doi: 10.1093/лесная наука/10.3.361
Полнотекстовая перекрестная ссылка | Академия Google
Жирнова Д. Ф., Белокопытова Л. В., Бабушкина Е. А., Кривелларо А., Ваганов Е. А. (2021). Структура ранней древесины вечнозеленых хвойных деревьев у границы леса определяется средой обитания, а поздняя древесина зависит от вида и времени года. Деревья 35, 479–492.doi: 10.1007/s00468-020-02050-2
Полнотекстовая перекрестная ссылка | Академия Google
Жирнова Д.Ф., Белокопытова Л.В., Барабанцова А. Е., Бабушкина Е.А., Ваганов Е.А. (2020). Что преобладает в климатической реакции Pinus sylvestris между границами ее ареала в горах: экспозиция склона или возвышенность? Междунар. Дж. Биометеорол . 64, 333–344. doi: 10.1007/s00484-019-01811-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зиако, Э., Бионди Ф. и Генрих И. (2016). Ячеистая дендроклиматология древесины: тестирование новых заменителей на сосне щетинистой Большого Бассейна. Фронт. Завод Наука . 7:1602. doi: 10.3389/fpls.2016.01602
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Река Енисей Центральной Сибири в позднем плейстоцене на JSTOR
Абстрактный Недавние палеолитические работы вдоль среднего течения реки Енисей в Центральной Сибири выявили долгую историю заселения, которая почти наверняка начинается в среднем плейстоцене.Хотя свидетельства нижнего палеолита несколько предварительны, есть веские основания полагать, что охотники-собиратели периодически занимали средний Енисей до последнего межледниковья. Степная среда региона в верхнем плейстоцене была относительно богатой; было обнаружено более 200 стоянок верхнего палеолита как до, так и после последнего ледникового максимума. Похоже, что этот регион был заброшен во время последнего ледникового максимума. Большая часть советских и российских археологических работ была ориентирована на культурно-историческую направленность, но в последнее время возрос интерес к разработке адаптационистских и экологических исследовательских стратегий.Средний Енисей и более широкий регион Центральной Сибири являются ключом к пониманию ранней адаптации к северу и размеров палеолитических перемещений населения.
Степная среда региона в верхнем плейстоцене была относительно богатой; было обнаружено более 200 стоянок верхнего палеолита как до, так и после последнего ледникового максимума. Похоже, что этот регион был заброшен во время последнего ледникового максимума. Большая часть советских и российских археологических работ была ориентирована на культурно-историческую направленность, но в последнее время возрос интерес к разработке адаптационистских и экологических исследовательских стратегий.Средний Енисей и более широкий регион Центральной Сибири являются ключом к пониманию ранней адаптации к северу и размеров палеолитических перемещений населения.
Журнал археологических исследований объединяет самые последние обзоры международных исследований по широкому кругу тем и географических областей. Этот авторитетный обзорный журнал улучшает доступ к растущему объему информации и литературы за счет публикации оригинальных критических статей, каждая в формате 25-40 страниц. Рецензируемые современные исследования по выбранной теме охватывают важные полевые исследования и открытия, а также обзор недавно опубликованной литературы в избранной области.
Рецензируемые современные исследования по выбранной теме охватывают важные полевые исследования и открытия, а также обзор недавно опубликованной литературы в избранной области.
Springer — одно из ведущих международных научных издательств, выпускающее более 1200 журналов и более 3000 новых книг ежегодно, охватывающих широкий круг предметов, включая биомедицину и науки о жизни, клиническую медицину, физика, инженерия, математика, информатика и экономика.
.
