Как сегодня клюет карась
ДАЛЕЕ…
Почему у других клюет, а у тебя нет? КАК СЕГОДНЯ КЛЮЕТ КАРАСЬ. Улетный клев обеспечен
здесь вообще рыба не водится». Плюсы и Минусы. Сегодня — ВоскресеньеПоложительные факторы: Благоприятная фаза луны для Карася: хороший клев Карася Оптимальное давление для клева карася.
» Здесь уже не до построения планов, на первый взгляд, что лучше всего караси клюют в тех водоемах, реже топя его или уводя в. Самое читаемое сегодня. Обычно карась поднимался, зачастую объясняется е Календарь кл ва карася когда клю т карась. Представляем к вашему вниманию календарь кл ва карася по месяцам. В этом рыболовном календаре содержится общая информация о кл ве карасей в водо мах России. Загадки Кл ва Карася. В случае, осенью-
Представляем к вашему вниманию календарь кл ва карася по месяцам. В этом рыболовном календаре содержится общая информация о кл ве карасей в водо мах России. Загадки Кл ва Карася. В случае, осенью- 
Ночной клев. Куда челябинским рыбакам отправиться за карасями для новогоднего пирога
На носу Новый год, но многие из нас так еще и не успели выбраться на рыбалку, ибо то рабочие будни время съедали, то семейные хлопоты.
 Так куда же отправиться его половить?!
Так куда же отправиться его половить?!Подкормить лунки
Биликуль — озеро настолько известное в рыболовной среде Челябинской области, что о его местонахождении говорить не приходится. Туда даже слепой за рулем доберется без всяких хлопот. Курганский тракт, Миасское — Тирикуль, еще с десяток километров, и вот оно, озеро с карасями, средний вес которых 180-300 граммов. Вкуснятинка, да и только, особенно в духовке под майонезом. А какие из этой рыбки шикарные пироги! Однако некоторые члены нашей рыболовной среды поглядывают на это озеро с легким подозрением, мол, радиация, и все такое… Напрасно! Все это пустопорожние слухи. Проверено на все сто!
Карась на Биликуле берет и днем, но лучше ночью. Видимо, небольшая глубина этого водоема (до 3 метров) в светлое время суток делает местных обитателей осторожными. Зато ночью, если вам повезло и вы оказались на рыбной дороге, вторая удочка становится неуместной. Двухсот- (и более) граммовые рыбы просто цепляются одна за другую.
Уважаемые рыбачки! Не стоит раскатывать губу на 120 верст (именно столько до села Биликуль, которое находится на побережье одноименного водоема). Карася здесь много, но все же это не базарный рыбный прилавок, где можно запросто отовариться, не прилагая никаких сил. Многие рыболовы, уповая на легкий успех, убывали отсюда несолоно хлебавши. Спросите почему? Ответ прост: все, как всегда, зависит только от вас. Если вы на водоеме, не утруждая себя поиском рыбы, упали где попало, можете не сомневаться: останетесь без улова.
А вообще, поймать 15 килограммов карася на Биликуле за ночь не так уж и сложно. Нужно просто засветло подсуетиться с поиском рыбы — побегать по озеру, то есть обойти дневных рыболовов с одним простым вопросом: ну, как дела? Разумеется, большинство из них пошлет вас по известному адресу, ибо никому не нужны соседи под боком, но некоторые промышляющие на льду окажутся душевными братьями и обязательно поделятся заветной информацией о том, что клюет. И если за день с лунки был пойман десяток-другой карасишек, то на этом месте можно смело ставить палатку для ночной рыбалки. Понятно, что заниматься подобным опросом не очень спортивно, зато экономия времени и прикормки здесь налицо.
Понятно, что заниматься подобным опросом не очень спортивно, зато экономия времени и прикормки здесь налицо.
Бутерброд с мотылем
Напомним, карась — рыба из семейства карповых, а посему морозная погода не для него. Если на термометре минус 20 градусов, отправляться на рыбалку — немалое заблуждение, хотя без исключений и сюрпризов иногда не обходится. В прошлом по самому первому льду эту рыбу мы успешно ловили даже в очень холодные дни, но, как известно, первый лед на то он и первый, чтобы сюрпризы преподносить.
Как правило, наибольшее оживление карася бывает в многодневные оттепели, с температурой около нуля градусов. Именно в такую погоду, когда на дворе слякоть или идет снег, карась активно начинает питаться и радовать своими поклевками рыбачков.
В зимний период карась в основном ловится на опарыш и мотыль или на то и другое одновременно (бутерброд). В ночные часы хороший результат дает подсадка к мотылю мелко нарезанного белка вареного яйца или сала.
 Именно подобное сочетание подчас бывает единственным ключиком к аппетиту рыбы, когда случается перелом погоды и рыба начинает хандрить. Прикормка — все тот же мотыль и сухой «Делфи», хотя некоторые рыболовы предпочитают магазинным прикормкам обычные домашние сухари с жареными семечками, пропущенные через мясорубку. Такая прикормка довольно быстро привлекает даже пассивную рыбу, создавая в воде запах и муть.
Именно подобное сочетание подчас бывает единственным ключиком к аппетиту рыбы, когда случается перелом погоды и рыба начинает хандрить. Прикормка — все тот же мотыль и сухой «Делфи», хотя некоторые рыболовы предпочитают магазинным прикормкам обычные домашние сухари с жареными семечками, пропущенные через мясорубку. Такая прикормка довольно быстро привлекает даже пассивную рыбу, создавая в воде запах и муть.Ночная рыбалка — дело весьма утомительное, поэтому обставлять ее нужно как можно с большим комфортом. И по наш день в XXI веке некоторые начинающие рыболовы до сих пор щурятся на поплавки, сидя в палатках с обычными свечками. При этом носы рыболовов краснеют от холода, а лунки покрываются льдом. Романтика! Как при этом им удается еще что-то поймать, вызывает лишь удивление. Мы в качестве отопления используем газ, а благодаря компактному аккумулятору и светодиодной лампе в палатке всегда светло будто днем. От рыбалки нужно получать удовольствие, а не ангину или какой-нибудь гайморит.
Крючки по-летнему
Итак, рыбка найдена, палатка поставлена. Прикормка: как будем кормить? Здесь приемы у всех свои, хотя особого разнообразия нет. В большинстве случаев это постоянное подбрасывание небольших порций мотыля и «пыление» размоченным сухарем. Интенсивность этих действий зависит от количества рыбы, появляющейся под вашей лункой: чем больше рыбы, тем чаще прикормка, но все же перекармливать никогда нельзя. До января карась активно передвигается по определенному участку водоема, поэтому поклевки происходят как бы с наскока: ловим подряд 3-6 рыбин, затем 15-минутная пауза. Именно в этот временной промежуток следует попылить сухариками и подбросить мотыль. Некоторые рыболовы, действующие по принципу «побольше накормлю — побольше наловлю», зачастую остаются с носом. Только дозированность прикормки дает ожидаемый результат.
Карась не слишком разборчивая рыба, поэтому оснастка удочки не требует каких-либо особых тонкостей. Для ловли карася зимой, как правило, используется леска диаметром 0,12-0,14 мм и крючки примерно 5-го номера, подвязанные на «летний» манер: крючок — груз — крючок. В простонародье такая оснастка называется «крестьянкой» и считается одной из самых уловистых для этого вида рыб. Но больший эффект придают ловле карася не простые крючки, а фосфорные мормышки, которые слегка мерцают в воде. В предыдущие ночные посиделки основной улов из 25 килограммов был пойман как раз на них, причем чаще клевало на мормышку вверху, подвязанную на поводке. В январе карась будет менее подвижен и наверняка перейдет на кормление со дна.
Клевые озера
Подведем итог. Ночная рыбалка на озере Биликуль сегодня остается одной из самых увлекательных и результативных. Но! Здесь, как и на других водоемах, нельзя полагаться на русский авось. Чтобы быть с богатым уловом, на этом озере рыбку нужно искать! И только тогда будет вам рыбацкое счастье и карасевые пироги!
На втором месте по карасевой популярности находится озеро Феклино, которое расположено в том же районе и до которого также около 120 верст. Маршрут примерно все тот же: Челябинск — Миасское — Бродокалмак — Алабуга — Феклино. Карась здесь помельче, 100-150 граммов, но поймать его проще: этой рыбы здесь пруд пруди. Понятное дело, когда рыбка клюет, то количество наезжающих сюда рыболовов становится запредельным, что и происходит на сегодняшний день. Принцип ловли такой же, как на Биликуле, насадка и прикормки те же — сухарь и мотыль.
Карась здесь помельче, 100-150 граммов, но поймать его проще: этой рыбы здесь пруд пруди. Понятное дело, когда рыбка клюет, то количество наезжающих сюда рыболовов становится запредельным, что и происходит на сегодняшний день. Принцип ловли такой же, как на Биликуле, насадка и прикормки те же — сухарь и мотыль.
Следующий часто посещаемый нашими челябинцами водоем — это озеро Чистое, где карась несколько крупнее феклинского, а следовательно, пожирней в пирогах. Находится Чистое уже в Курганской области, но всего в 70 верстах от нашего города, недалече от станции Пивкино, и если заглянуть в карты «Гугл», то его легко будет найти. Принцип ловли «чистого» карася тот же, как и на вышеперечисленных озерах, к которым можно добавить еще такие карасевые водоемы, как Пуктыш, Айбатово, Алабуга, Линева, Таволжаное… В общем, было бы желание, а нам, южноуральцам, есть куда поехать за карасем.
Мы поздравляем всех рыболовов с наступающим 2018 годом, желаем всего хорошего — и ни чешуи ни хвоста! До новых встреч в новом году!
На какой глубине ловить карася в разные сезоны
Карась – рыба всем известная и многими любимая. Некоторым кажется, что карасевая рыбалка – максимально простая и понятная, и никаких особых вопросов в ней нет – надо только насадить на крючок червя и закинуть куда-нибудь оснастку. Но на самом деле, конечно, в ловле карася есть довольно много нюансов, в которых надо разбираться, чтобы получать хорошие уловы в самых разных условиях. И один из важных моментов – выбор глубины для рыбалки.
Некоторым кажется, что карасевая рыбалка – максимально простая и понятная, и никаких особых вопросов в ней нет – надо только насадить на крючок червя и закинуть куда-нибудь оснастку. Но на самом деле, конечно, в ловле карася есть довольно много нюансов, в которых надо разбираться, чтобы получать хорошие уловы в самых разных условиях. И один из важных моментов – выбор глубины для рыбалки.
Какое дно предпочитает карась
Охотнее всего эта рыба держится на илистом дне. Копаясь в нем, она легко находит себе пропитание – к примеру, мотыля и других личинок насекомых. Недостатков у заиленных участков два:
- Для обитателей водоемов минус в том, что в таких местах довольно мало кислорода – гораздо меньше, чем на песчаном или каменистом дне. Но для карася это большого значения не имеет. К содержанию кислорода в воде эта рыба совсем не требовательна.
- Второй недостаток ловли на заиленном дне, который имеет значение уже только для рыбака – оснастка с приманкой может легко в нем «утонуть», став недоступной для рыбы.
 Но избежать этого несложно, надо только использовать правильные монтажи.
Но избежать этого несложно, надо только использовать правильные монтажи.
 Но избежать этого несложно, надо только использовать правильные монтажи.
Но избежать этого несложно, надо только использовать правильные монтажи.Таким образом, именно на заиленных участках и стоит искать карасика в первую очередь. Ловля здесь приносит наилучшие результаты.
Карасевая рыбалка часто проходит и на песчаном дне. Такие участки эта рыба тоже выбирает довольно охотно. Но по привлекательности для карася песчаное дно уступает илистому. Зато и ловить здесь проще. Крючок с приманкой гарантированно не уйдет в грунт, независимо от того, какая оснастка используется.
На каменистом дне встретить карася можно нечасто. Здесь эта рыба – редкий гость и долго не задерживается. На каменистых участках целенаправленно ее не ловят.
В этом видео рассказывается о том, где искать карася в реке, об особенностях мест его обитания и циклах миграций:
На какой глубине ловить карася в зависимости от сезона
В период с весны по осень поведение карася и участки, которые он выбирает для кормежки, довольно сильно меняются.
Весна
Весна – время года, в которое эта рыба больше всего перемещается по водоему, меняя и глубину стоянок.
До нереста
В первой половине весны, до нереста, вода еще холодная. Поэтому карась выходит кормиться на мелководья. На участках с минимальной глубиной вода хорошо прогревается солнцем. Здесь максимально оживляются разные донные беспозвоночные. Приходит сюда кормиться и рыба, самая разная – от небольших мальков до хищников, которые охотятся на других чешуйчатых.
Карась – не исключение. В первой половине весны, до нереста, он охотно кормится на самых небольших глубинах – от 0,5 м, 1 м, 1,5 или 2 м. Именно здесь его и стоит искать в первую очередь. На таких мелководьях всегда можно встретить довольно много активного карася, который вовсю ищет пропитание и поэтому один за другим попадается на крючок. На ямах в это же время остается та рыба, которая еще не кормится или кормится мало.
Внимание! Крупный карась почти всегда предпочитает держаться на глубине. Это справедливо и для весны. Если вы хотите половить именно трофейные экземпляры, в это время их нужно искать на глубинах как минимум от 1 до 2 м или скорее от 1,5 до 2 м.
Это справедливо и для весны. Если вы хотите половить именно трофейные экземпляры, в это время их нужно искать на глубинах как минимум от 1 до 2 м или скорее от 1,5 до 2 м.После нереста
После икромета карась как бы разбредается по водоему. Успешно ловить его в это время можно и на малых, и на средних, и на больших глубинах. Если вас интересует трофейная рыбалка, надо искать рыбу на ямах. Средние и мелкие экземпляры кормятся под берегом и на небольшом удалении от него, то есть на малой и средней глубине.
Лето
В летнюю пору карась, в зависимости от ситуации, может хорошо ловиться на небольших, средних или больших глубинах.
На ямы рыба уходит кормиться в жару, в разгар лета. На небольших и средних глубинах в это время вода слишком сильно прогревается солнцем, и ей становится некомфортно находиться здесь.
Внимание! Если жара длится долго, неделю, две и больше, и температура воды поднимается слишком высоко даже на ямах, рыба поднимается к поверхности «подышать воздухом». Но при этом она не кормится, и ловить ее бесполезно.
Но при этом она не кормится, и ловить ее бесполезно.
Участки с небольшой глубиной в летний период – хороший вариант для ночной ловли карася. В темноте рыба подходит кормиться к самому к берегу, и можно получить отличные уловы, используя самую простую легкую маховую удочку. Причем среди трофеев могут быть экземпляры весом в полкилограмма, килограмм и даже больше. Правда, на разных водоемах в различных регионах ситуации сильно различаются, и такая добычливая ночная рыбалка есть далеко не везде.
Средняя глубина – это «универсальный вариант» для летней рыбалки. Здесь карасик попадается в течение всего лета, как в теплую погоду, так и в прохладную, это может быть и небольшая рыба, и хорошие крупные экземпляры.
Осень
С окончанием лета карась уходит от берегов подальше, на максимальную глубину. Именно здесь его и надо искать осенью. Рыба нагуливает жир и готовится к зимовке, которая пройдет здесь же, на ямах. Искать ее на небольших глубинах под берегом можно только в начале осени.
В это время карась еще во многом придерживается летних привычек. В погожие солнечные дни он может выходить на мелководья, где вода хорошо прогревается. Но ловить его в таких местах во второй половине осени – совершенно бесперспективное занятия.
Глубина для ловли карася в зависимости от водоема
Многое зависит от характера водоема и его особенностей. Так, на мелководных прудах и озерах (с глубинами до 2 м) карася надо искать у дна. В средних и верхних слоях воды ловить нечего. На таких водоемах глубину отпуска приманки лучше выбирать всегда максимальную, за исключением разве что весеннего периода.
Если на дне водоема бьют холодные ключи, и температура воды в придонном слое меньше, эта рыба обычно держится на границе холодного и теплого слоев либо выше, уже в хорошо прогретом горизонте.
Выбирать глубину ловли карася в таких озерах и реках довольно сложно. Нужно либо использовать хороший эхолот или видеокамеру, которые смогут показать, на каком горизонте находится рыба, либо подбирать этот горизонт экспериментально, делая пробные забросы с разным отпуском приманки.
Если в водоеме есть пищевые конкуренты карася – обычно это карп и линь – он часто предпочитает держаться повыше, в средних слоях воды или у поверхности. На таких прудах и озерах нередко приходится выставлять оснастку так, чтобы приманка погружалась на глубину в 10-20 см и не больше. Подобный подход приносит здесь лучшие уловы этой рыбы.
Таким образом, выбор глубины ловли карася – не такая простая задача, как может показаться на первый взгляд. Иногда эта рыба держится под самым берегом, на мелководье, а иногда, наоборот, уходит ближе к середине водоема. Также карась может выбирать разные горизонты – искать пропитание у дна, в средних слоях или под самой поверхностью. Но, если знать основные правила выбора глубины для его ловли и больше экспериментировать, подавая оснастку в разные точки и устанавливая разный отпуск приманки – успех гарантирован.
Полезное видео
В этом видео рассказывается о 5 основных способах ловли карася:
 youtube.com/embed/49D4NUM0wUY» frameborder=»0″ allowfullscreen=»allowfullscreen»/>
youtube.com/embed/49D4NUM0wUY» frameborder=»0″ allowfullscreen=»allowfullscreen»/>
Ловля карася в прудах
Караси бывают двух видов — золотой и серебряный. Золотой обычно встречается в мелководных, илистых, заросших водными растениями, особенно рдестом, водоемах. Серебряный распространен более широко. В больших современных прудах он потеснил своего собрата так основательно, что об отдельной ловле золотого карася говорить не приходиться.
В нашей стране известны поимки особей золотого карася 45 см длиной и весом до 3 кг, а самый крупный зарегистрированный за последние годы экземпляр серебряного карася весил лишь чуть более двух килограммов.
Несмотря на это, картина в целом на наших прудах обратная. Серебряный карась на некоторых водоемах ловится нередко и до килограмма весом, а золотой карась больше известен как карликовая форма в старых деревенских запрудах.
Каждый, кто хотя бы несколько зорь провел за ужением этой рыбы, может подтвердить сказанные около ста лет назад слова: «Сегодня он берет отлично, завтра совершенно не клюет, и трудно объяснить почему».
Так отозвался в свое время о карасе известный русский рыболов и писатель Л.П. Сабанеев. Но и сегодня нередко можно встретить опытного рыболова, вернувшегося с ловли карася «пустым». Нет, не зря говорят: «Чтобы быть удачливым карасятником, стань сначала страстным поклонником этой рыбы».
Еще лет двадцать назад карась легко покорялся тому, кто постоянно прикармливал удачливое место. Сегодня такая тактика может дать осечку. Нынешние пруды становятся непредсказуемыми по причинам, чаще всего связанным с сельскохозяйственной химизацией.
Бывает, что один год пруд дает отличный улов, а на другой его словно подменили. Поэтому на протяжении всего лета нужно иметь самые свежие данные о клеве карася на различных водоемах. Полезно знать все: как вдет лов, на какие насадки, снасти.
Нерестится карась на четвертом-пятом году жизни. На широте Марийской республики нерест проходит в конце мая — начале июня и совпадает с цветением калины. Устоявшееся время нереста подтверждает и старинная марийская примета — «полан пеледеш — карака модеш» (цветет калина — играет карась). Впрочем, в иные годы приходится наблюдать нерест карася дважды и даже трижды. Такое случается в прохладное, дождливое лето, когда температура воды непостоянная. Этим, пожалуй, можно объяснить и такую марийскую примету, записанную А. Китиковым: «Карака кум гана модеш: ломбо да полан пеледме годым, шульо сорла годым» (карась играет трижды: во время цветения черемухи, калины и уборки овса).
Впрочем, в иные годы приходится наблюдать нерест карася дважды и даже трижды. Такое случается в прохладное, дождливое лето, когда температура воды непостоянная. Этим, пожалуй, можно объяснить и такую марийскую примету, записанную А. Китиковым: «Карака кум гана модеш: ломбо да полан пеледме годым, шульо сорла годым» (карась играет трижды: во время цветения черемухи, калины и уборки овса).
Определить близость нереста можно и не полагаясь на приметы. Для этого сгодится обычный термометр. Стоит только запомнить, что карась начинает икромет при температуре воды + 16—18° С.
Изредка и под осень можно поймать карася, не отметавшего икру. Если это связывать с погодой, то будет неверно. Результаты последних исследований ученых говорят о том, что у карася очень мало самцов. Обычно их не более 5—30 процентов от общего количества рыбьего стада.
Такое соотношение связано с тем, что у данной рыбы необычная форма размножения — гиногенез. Поэтому нужно скептически отнестись к распространенному представлению о серебряном карасе как результате скрещивания золотого карася с карпом. При гиногенезе спермин самца не участвуют в развитии яйцеклеток, а только активизируют их. Лишь в Сибири карась порой встречается с обычным типом осеменения, то есть двуполый, как и остальные наши пресноводные рыбы.
При гиногенезе спермин самца не участвуют в развитии яйцеклеток, а только активизируют их. Лишь в Сибири карась порой встречается с обычным типом осеменения, то есть двуполый, как и остальные наши пресноводные рыбы.
Ранней весной, когда вода прогреется до 8°С, карась выходит на мелководье и начинает ловиться на удочку. Если вода еще мутна, то ловить можно у самого берега. Короткое, до 1,5 метра, удилище, леска 0,15—0,17, крючок №2,5—5 и самые разнообразные, по вкусу, поплавки — вот снасть, которая принесет успех в это время и начинающему рыболову.
Со временем вода прогревается, и карась начинает ловиться на более глубоких местах. Однако туда не стоит пока особенно стремиться. До начала нереста карась с успехом клюет на глубине от 20 до 50 см. А если у вас круглый, неброско окрашенный небольшой поплавок, то не стесняйтесь испытать снасть там, где глубина всего сантиметров пятнадцать. Нужно только, чтобы это было возле торчащей из воды коряжины, куста или на месте затопленной прошлогодней травы.
Порой диву даешься, каких крупных карасей ловят на мелководье! Высота их тела может лишь на несколько сантиметров быть меньше глубины затопленного места. Вода после подсечки в таких случаях словно взрывается и вместо танцующего поплавка внезапно видишь упругое тело рыбы.
Цветение шиповника совпадает с началом лучшего донерестового клева. Если в эти дни встанет проблема выбора насадки, то прежде всего остановите свой выбор на мелком навозном черве.
Но насаживать его нужно не пучком, как на реках, а по одному, «чулочком». Если червь велик, то обрывайте головку. Хвостик на крючке дольше шевелится, особенно, если оставите свободным его последний сантиметр. Кстати, нередко в это время карась берет и на затрепанный обрывок червя. Когда клев исключительно хорош, то на такой оборвыш можно без проблем поймать несколько рыб!
Преднерестовый клев карася — время, когда почти из любого человека можно сделать заядлого рыболова.
О ловле рыбы во время нереста обычно не пишут. И относительно большинства рыб на такую ловлю существуют запреты и ограничения. Но карась фактически не отнесен к ценным рыбам и обитает в водоемах, где ловля удочкой не запрещена круглый год. Плюс к тому — клев карася в его нерест возможен.
И относительно большинства рыб на такую ловлю существуют запреты и ограничения. Но карась фактически не отнесен к ценным рыбам и обитает в водоемах, где ловля удочкой не запрещена круглый год. Плюс к тому — клев карася в его нерест возможен.
Выскажу только несколько практических советов.
Во время нереста не рассчитывайте на большой улов. Помните, что первыми икромет начинают крупные особи, затем — мелкие. Соответственно и клюют сначала мелкие караси, а затем, отдохнув после нереста, более крупные. Лучшей насадкой считаю опарыш.
Максимально постарайтесь использовать этот период для испытания различных новшеств в оснастке. Кроме того, узнайте, как обстоят дела на соседних водоемах. На новых полюбившихся местах исследуйте характер дна, возможность зарастания их водными растениями. Постарайтесь затем проанализировать, почему рыба держится в данном месте? Нелишне и благоустроиться: убрать мешающие ловле нависшие ветви деревьев, определить постоянное место для костра и ночевок.
Вскоре после нереста можно начать приваживание. Желательно, чтобы это происходило регулярно и в одно и то же время дня. Место приваживания очищают от растительности и делают «стол» из высыпанного на дно одного-двух ведер речного песка.
После нереста случается кратковременное бесклевье. Особенно оно заметно после короткого, но дружного нереста. Затем вновь недели две клев отличный, почти как перед нерестом. Насадки самые разнообразные. Но на этом золотая пора для рыболовов кончается. Далее идет время, не поддающееся устойчивым рыбацким прогнозам. Во многом все будет зависеть как от погодной характеристики лета, так и от изменений в кормовой базе каждого отдельного водоема. Больше повезет, конечно же, тем, кто имеет возможность постоянного прикармливания рыбы. Но таких рыболовов меньшинство.
Если же брать более короткие отрезки времени, скажем, неделю, то здесь непредсказуемого еще больше. Хотя некоторые «правила» все же существуют.
Например, в небольших прудах, где карась — основная рыба, клев нередко происходит волнообразно. То есть после одного-двух дней отличного клева идет постепенный его спад. Это не касается лишь водоемов, где этой рыбы изобилие, но она карликовой формы.
То есть после одного-двух дней отличного клева идет постепенный его спад. Это не касается лишь водоемов, где этой рыбы изобилие, но она карликовой формы.
Пасмурная погода с теплым дождиком чаще всего не помеха. В такие дни карась может ловиться с утра до вечера. А вот в жаркие безветренные дни, когда великолепны утренние зорьки, клева можно вообще не увидеть. И спасает только ночная ловля.
Если говорить об отдельно взятом дне, то летом клев карася лучше утром, нежели вечером. И начинать лов можно с самого рассвета, едва станет видно поплавок. В эти первые минуты и с 4 до 5 часов утра, а также ночью, нередко берут самые крупные экземпляры.
Значительно улучшает клев прошедший дождь.
Бывают дни, когда лов с берега ухудшается в течение дня. В таком случае выручает ловля с лодки. Но чтобы вовремя сменить ее способ и предугадать отход карасей с мелководья, нужно следить за атмосферным давлением. Колебания его — верный признак перемещения рыбы, что заядлыми береговыми рыболовами постоянно принимается за прекращение клева вообще.
Летним вечером карась начинает брать особенно хорошо где-то за два часа до захода солнца и ловится до глубоких сумерек.
Есть любители и ночной ловли. Ими не без оснований самой удобной снастью считается донка с резиновым амортизатором (к ней еще вернемся ниже). Но ночью ловля все же малоприятна и более утомительна, поэтому поклонников у нее гораздо меньше. На ночную ловлю идут обычно тогда, когда в водоеме есть крупная рыба, и она не хочет ловиться в иное время суток. А также в том случае, когда целый день на водоеме находятся купальщики.
Ночью можно ловить и на поплавочную удочку, используя достаточно мощный фонарик. Лучшее время ловли — с наступления темноты до полуночи, затем, после одно-двухчасового перерыва клев возобновляется. Иногда ночной улов весомее утреннего.
Современные пруды, созданные с мелиоративной целью, имеют солидные размеры. И тем самым, на мой взгляд, внесли в ловлю существенные изменения.
Рядом с карасем здесь стали сосуществовать различные рыбы: окунь, плотва, пескарь и даже щука. И в некоторых подобных водоемах карася серьезно потеснила хищная рыба. Однако там, где немало коряжистого мелководья и густой водной растительности, после нескольких лет жесткой борьбы за выживание карась сохранился в большом количестве. Это крепкие, в сто и более граммов весом, «серебряные» рыбы охотно клюют и приносят немало радости рыболовам, тем самым опровергнув давнишнее утверждение многих авторов рыболовных справочников, гласящее о том, что хороший клев происходит только в чисто карасевых прудах и озерах, где нет другой рыбы.
И в некоторых подобных водоемах карася серьезно потеснила хищная рыба. Однако там, где немало коряжистого мелководья и густой водной растительности, после нескольких лет жесткой борьбы за выживание карась сохранился в большом количестве. Это крепкие, в сто и более граммов весом, «серебряные» рыбы охотно клюют и приносят немало радости рыболовам, тем самым опровергнув давнишнее утверждение многих авторов рыболовных справочников, гласящее о том, что хороший клев происходит только в чисто карасевых прудах и озерах, где нет другой рыбы.
Ловить в больших прудах, где «победил» карась, одно удовольствие. Но и здесь можно остаться без улова, если забыть давнишнюю особенность карася — любовь к частому изменению своего «меню».
Питается карась довольно разнообразным кормом: рачками, моллюсками, червями, личинками насекомых, в том числе и мотылем. Кое-где, например, в Алтайском крае, на некоторых прудах ловят эту мирную рыбу и на малька, и на рыбье мясо. Но это исключения, говорящие больше о том, что с кормовой базой водоема не все в порядке. Основным же кормом, по утверждению ученых, для карася является мягкая водная растительность. В этом можно убедиться и самому, вскрывая пойманную рыбу. Кишечник ее зачастую зеленого цвета от проглоченной растительной пищи.
Основным же кормом, по утверждению ученых, для карася является мягкая водная растительность. В этом можно убедиться и самому, вскрывая пойманную рыбу. Кишечник ее зачастую зеленого цвета от проглоченной растительной пищи.
Вполне вероятно, что именно в этом корме кроются разгадки знаменитого карасиного непостоянства. Но в литературе пока не прочитать, а от рыболовов не услышать об удачно примененной насадке из водной растительности.
Зато если вести речь о других насадках и способах их приготовления, то здесь самоё богатейшее разнообразие. Ни одна другая рыба не удостоилась от рыбацкого роду-племени такой массы рецептур приготовляемых «блюд», как карась.
Часть рецептов вы найдете в специальном разделе этой книги. Пока же отметим, что среди самых популярных насадок пальму первенства держит обыкновенный навозный червяк, длиной в 3—5 см. Затем почти всегда называют опарыша (личинку синей мясной мухи), хотя в некоторых местностях его упорно избегают вообще — традиция! Затем не менее популярны тесто и белый хлеб. В последние годы стали упоминать и черный хлеб. Не удивляйтесь, но если сюда добавить популярную также манную кашу, то получится только минимум, что благоразумнее всего взять с собой, шагая на незнакомый водоем.
В последние годы стали упоминать и черный хлеб. Не удивляйтесь, но если сюда добавить популярную также манную кашу, то получится только минимум, что благоразумнее всего взять с собой, шагая на незнакомый водоем.
Применение насадок при ловле карася также своеобразно. Ранним утром он может хорошо ловиться на опарыша, после восхода солнца постепенно увлечься червем или соблазниться шариком хлеба, а вечером вновь изменить свои пристрастия.
Бывают дни, когда капризность этой рыбы потрясает и опытного рыболова. Весной карась иногда начинает брать лишь на хвостик червя. Причем на помятый и потерявший всякий вид. Летом — другой пример: берет на шарик белого хлеба, но неплотно скатанный и лишь от сдобной булки. И не тронет насадку из обычного хлеба, будь она и медом смазана.
Особенно такая привередливость встречается, по моему наблюдению, на небольших прудах, где у карася нет серьезных конкурентов в добыче пищи и нет хищной рыбы.
На больших прудах, где рыбное семейство разнообразно, чаще иная сложность — правильно подобрать место ужения.
В небольших, площадью до десяти гектаров, прудах удачным местом будет почти всегда то, которое вам понравится. Исключение из правил — места у самого водосброса и заболоченный конец пруда. А утоптанная предыдущими рыболовами площадка у воды улов почти гарантирует.
В более крупных водоемах нужно исходить из наличия на них водных растений, расположения глубин и отмелей, затопленных кустов и т. п.
Где берега пруда голые, а на поверхности воды нет или почти нет водной растительности, нужно попробовать начать ловлю в местах с изменениями в береговой линии, что предполагает и изменение рельефа дна. Это могут быть небольшие мыски и заливчики, края песчаных отмелей (рис.1). Если есть возможность половить несколько дней, то лучше всего попробовать такой хорошо зарекомендовавший себя способ привлечения рыбы, как «искусственный» коряжник. Охапка притопленных сухих лапок ельника через пару дней станет излюбленным местом посещения карасей.
Рис 1.Если на водоеме имеется небольшой пляж с твердым или даже песчаным дном, то здесь возможен успех ночью, при ловле на донку. Утром и вечером предпочтительнее места на подходе к пляжу. Ловля днем также возможна, но для этого нужно отойти на мелководье, к началу пруда, где нет купальщиков и заиленное дно. Кстати, группа больших деревьев или лесок за спиной в этом случае не помешают. Помогает маскироваться и легкая рябь на воде.
Утром и вечером предпочтительнее места на подходе к пляжу. Ловля днем также возможна, но для этого нужно отойти на мелководье, к началу пруда, где нет купальщиков и заиленное дно. Кстати, группа больших деревьев или лесок за спиной в этом случае не помешают. Помогает маскироваться и легкая рябь на воде.
При ловле днем не нужно бояться ветра. Если он не особенно сильный и не порывист, то это только на пользу делу. Имея легкую оснастку, можно половить в проводку. Перяной поплавок, высоко торчащий из воды, или приподнятая в воздух леска послужат своего рода парусом. Насадка при движении снасти почти задевает дно. Такая проводка может принести неплохой результат.
В водоемах, где имеется водная растительность, карася нужно искать прежде всего возле нее. И здесь, в отличие от рек, рыба зачастую не избегает водорослей роголистника и элодеи. Видимо, это связано с тем, что иной растительности зачастую на прудах не бывает. А защиту и корм карасю искать приходится.
Некоторые рыболовы утверждают, что карасиные тропы проходят вдоль кромки водяных зарослей. Это, считаю, не совсем так.
Это, считаю, не совсем так.
Вооруженный удочкой рыболов ищет удобные для ловли места, а заросли водных растений для этого менее всего пригодны. Но стоит расчистить от водорослей «полянку» и прикормить ее, как карась тут же даст о себе знать. Следует только при этом помнить, что глубина в данном случае должна быть не менее 30 см. Объяснение простое — «шапка» водорослей на поверхности достигает значительной толщины, а карась, плавая под зарослями, любит относительную свободу.
Расчистка «полянки» в сплошных зарослях водорослей дает лучший результат, если до чистой воды не более полтора метра, а также, если рядом коряжник. Иногда достаточно и того, чтобы к месту ловли подходил конец лежащей на дне коряжины.
Если вам некогда заниматься расчисткой места, то присмотрите в водных зарослях заливчики чистой воды. Обычно они показывают на имеющееся углубление дна. И почти всегда это отличные места для лова.
В начале лета, когда водных зарослей на поверхности мало, на границе их с чистой водой клев хуже, чем отступя полметра-метр на глубину. Вполне возможно, что ростки новых растений больше привлекают карася, чем взрослые растения.
Вполне возможно, что ростки новых растений больше привлекают карася, чем взрослые растения.
Иногда в места ловли высыпают речной песок. Трудоемкость этого дела окупается не только хорошим клевом, но и отсутствием зацепов. Хорошо заметна на таком фоне и насадка.
Если же клев идет неплохо, то экспериментировать с целью улучшения места не всегда целесообразно. Значит, привлекательно это место само по себе. В случае же частых зацепов о водоросли на леске в сантиметре от крючка крепят кружок из тонкой пластмассы или засвеченной фотопленки. Темноватый цвет кружка не отпугнет рыбу, а близкое расстояние до крючка не позволит совершиться зацепу. Диаметр кружка тоже небольшой, около сантиметра.
Леску, идущую от удилища к поплавку, перед ловлей в водных зарослях лучше всего смазать жиром, парафином или даже лыжной мазью, так она не затонет и лишний раз не запутается.
Когда карася в водоеме много, и он клюет довольно устойчиво, то, выбрав нужную насадку, можно не вспоминать о приваде. Лишь иногда для активизации клева можно подбрасывать щепотку-другую прикормки, состоящей обычно из той же насадки, на которую в данный момент ловите.
Лишь иногда для активизации клева можно подбрасывать щепотку-другую прикормки, состоящей обычно из той же насадки, на которую в данный момент ловите.
Чем же воспользоваться, если без привады все же не обойтись?
Отличные результаты дает раздробленный и обваренный кипятком жмых. Достать его трудно, поэтому на замену нередко идет заваренный или просто замоченный комбикорм. Между тем поджаренные на подсолнечном масле семена конопли и подсолнечника прекрасно позволяют обойтись без жмыха. Нужно только растолочь их или пропустить через мясорубку, смешать с хлебом.
С привадой может выйти осечка, если пруд часто посещаем, и рыболовы не жалеют для рыбы каши и хлеба… Из-за гниющих остатков привады портится вода, да и рыба становится капризнее в выборе насадки. В таком случае помогают нетрадиционные способы приваживания — подброшенные к месту ловли подпаленные рога животных, пахучие добавки к насадке.
Из пахучих добавок обычны листья мяты, анисовые и валерьяновые капли, ваниль и т. д. Все они, хотя и часто упоминаются в рыболовной литературе, на деле редко становятся традиционными в арсенале рыболова. Причина в том, что не каждому в данном случае присуще чувство меры. Почему-то забывается, что карась, несмотря на свою неприхотливость, очень чувствительная к запахам рыба. И излишеств не любит.
д. Все они, хотя и часто упоминаются в рыболовной литературе, на деле редко становятся традиционными в арсенале рыболова. Причина в том, что не каждому в данном случае присуще чувство меры. Почему-то забывается, что карась, несмотря на свою неприхотливость, очень чувствительная к запахам рыба. И излишеств не любит.
Пора сказать о снастях.
Большую популярность приобрела ловля карася при помощи донки с резиновым амортизатором (рис. 2). Ею ловят как на илистом мелководье, так и на глубинах, а также в прогалах между водными растениями. Удобнее других снастей она при ловле ночью, так как снимается проблема точного заброса снасти.
Рис 2.Донка имеет около десяти крючков, довольно добычлива, поэтому долгое время находилась под запретом. Перенятая у волжских рыболовов, претерпела ряд изменений. Прежде всего ее не нужно постоянно держать в руках, шевелить насадку.
Оснастка проста. К тяжелому, весом не менее 300 граммов, грузу прикрепляют авиамодельную резину в 7—10 метров (редко длиннее), затем с помощью вертлюжка — леску. Сечение последней обычно 0,5 мм. Тоньше — чаще путается при выбирании снасти и режет пальцы, толще — менее чувствительна при поклевках. Поводки длиной от 10 до 20 см (в зависимости от удаленности от конечного груза) с крючками № 5—6 крепятся различно: узлом, «петля в петлю» или, что удобнее, через вертлюжки. Последние имеют по основной леске несколько сантиметров свободного хода, ограниченного стопорами из узлов.
Сечение последней обычно 0,5 мм. Тоньше — чаще путается при выбирании снасти и режет пальцы, толще — менее чувствительна при поклевках. Поводки длиной от 10 до 20 см (в зависимости от удаленности от конечного груза) с крючками № 5—6 крепятся различно: узлом, «петля в петлю» или, что удобнее, через вертлюжки. Последние имеют по основной леске несколько сантиметров свободного хода, ограниченного стопорами из узлов.
С помощью такой донки увеличивается возможность поимки крупной рыбы, не так осторожной вдали от берегов. Важно только, чтобы крючки касались грунта. А этого можно достичь при ровном дне и максимально отпущенной леске.
В бесклевье время от времени полезно делать небольшие, медленные потяжки с возвратом приманок на прежнее место или же сместив их на метр-два ближе к берегу. На большее смещение решиться рискованно — вследствие натяжения основной лески уменьшается чуткость снасти.
Вертикально закрепленный удильник на берегу обязательно должен иметь колокольчик (или заменяющий его отвес), который оттягивает леску почти до земли. Пусть колокольчик задевает при этом травинки, а это к лучшему. Меньше шансов ложного сигнала.
Пусть колокольчик задевает при этом травинки, а это к лучшему. Меньше шансов ложного сигнала.
Если же ветер силен и продолжает шевелить колокольчик (и даже отвес), то дальше под леску ставят одну, а то и две рогульки. Помогает и положенная на колокольчик легкая палочка.
Мелочи? Нет, это непременные условия для успешной ловли.
Саму поклевку определяют по сигналу колокольчика или движению отвеса. Сила потяжки, вопреки распространенному мнению, нередко от величины рыбы не зависит. Поклевка карася может быть настолько плавной, что колокольчики, которые вы приобрели в магазине, могут не сработать. Нужны очень чуткие колокольчики, которые лучше сделать самим из металлических охотничьих гильз.
Подсечка должна быть немедленной, еще до окончания потяжки. Быстрой, но не резкой.
Чтобы удобнее было снять пойманную рыбу, сзади удильника, на расстоянии, равном участку лески с крючками, крепят в земле прочный кол высотой метра полтора. Он будет держать за петлю на леске вынутую для проверки снасть, а крючки — на удобной для вас высоте. Ночью для ориентира и безопасности на кол лучше повесить белую тряпку. Она пригодится и для вытирания рук. А на земле возле кола удобно оставить и коробку с насадкой.
Ночью для ориентира и безопасности на кол лучше повесить белую тряпку. Она пригодится и для вытирания рук. А на земле возле кола удобно оставить и коробку с насадкой.
Для освещения снасти ночью лучше всего подходит фонарь типа «летучая мышь» или, что несколько хуже, крепящийся на головном уборе легкий фонарик. Применяют и свечу. А чтобы она была достаточно безопасна для лески и не задуваема ветром, ее крепят внутри стеклянной банки без дна или внутри растянутого на колышках целлофанового пакета с отрезанным дном.
Несмотря на то, что на эту снасть чаще берет крупная рыба, спортивного азарта при такой ловле явно не хватает. Одна из причин — сопротивление рыбы ощущается слабее, чем, скажем, на поплавочной снасти.
— Да и какая летняя ловля без поплавка? — удивится иной любитель карасиного ужения. И будет во многом прав. Ведь карась принадлежит к тем рыбам, поклевка которых отличается особенной красотой. Какому рыболову после дня ловли не снился медленно заваливающийся на бок «гусиный» поплавок?
Известный современный французский писатель и заядлый рыболов Аядре Стиль в своей повести «Рыбная ловля с пером» писал с сожалением, что синтетические изделия вытеснили со снастей рыболовов перяной поплавок. И на него сталo ловить во Франции просто стыдно.
И на него сталo ловить во Франции просто стыдно.
У нас, не знаю — надолго ли, дело обстоит пока иначе. Перяные поплавки можно встретить у рыболовов довольно часто. Бывают они и в продаже. В сочетании с леской 0,15—0,22, крючком № 3,5—5 и удилищем в 3—5 метров длиной, эти поплавки составляют типично «карасиную» удочку.
Многие правильно замечают, что для карася, как и для большинства мирных рыб, крючки предпочтительнее с прямым загибом, не из очень тонкой проволоки, остро заточенные. Цвет крючка особой роли не играет, хотя светлых нередко избегают, особенно, если ловят на червя. При ужении на растительные насадки и опарыш существует старое правило — использовать крючки с коротким цевьем.
Леску для карася нередко ставят излишней прочности, хотя достаточно сечения 0,15 мм. Другое дело, что за такой леской нужен более внимательный уход, о котором часто забывают. Но заменить ее в начале сезона и периодически смазывать (в том числе на зиму) глицерином не так уж и сложно. Тем более, что тонкая леска, с еще более тонким поводком, увеличивает количество поклевок в плохой клев. Придает ловле остроту ощущений, спортивность.
Придает ловле остроту ощущений, спортивность.
Под стать леске должно быть и удилище. Грубоватая вершинка не годится. Дело в том, что губы у карася довольно нежные и при сильной подсечке могут оборваться. Поэтому у «карасятников» не выходят из моды бамбуковые и даже рябиновые удилища. Хотя в последнее время их основательно теснят телескопические.
Техника ловли карася для многих представляется унылым сидением на месте. Это не так. Здесь масса тонкостей, которые трудно и перечислить. Но от них зависит величина улова даже в неплохой клев. Вот несколько примеров…
Попробуйте закинуть насадку на сантиметров 10—20 дальше обычного. А затем медленно подтащите ее к месту ловли. Если поблизости есть карась, то он не замедлит откликнуться на такое приглашение отобедать.
Можно действовать и по другому. С помощью удилища, если ловите недалеко от берега, несколько раз медленно поднимите и опустите поплавок. Или же, приподняв, мелко потрясите им так, словно ловите на мормышку. Кстати, она при ловле карася неплохо заменяет крючок, но изменяется характер поклевки, становится жестче, энергичнее.
Кстати, она при ловле карася неплохо заменяет крючок, но изменяется характер поклевки, становится жестче, энергичнее.
В литературе по ужению рыб нередки высказывания о том, что при ловле карася насадку нужно опускать на 3—5 см выше дна. Собственный опыт убедил меня в том, что лучший клев все же со дна.
Немаловажное значение при настрое снасти играет грузило. Имея в виду, что карась любит класть поплавок на бок, расстояние от крючка до грузила лучше всего оставлять равным примерно половине высоты тела предполагаемой рыбы. В этом случае (рис. 3) момент поклевки всегда своевременно отразится на поплавке.
Рис 3.Случается, что карась с осторожностью берет предлагаемую насадку и засекается только после скольжения в сторону положенного на поверхность воды поплавка. Заметив такую особенность, следует отодвинуть грузило к поплавку на 10—12 см. В этом случае поплавок среагирует лишь на главный момент — потяжку поплавка в сторону или вглубь.
Речь в обоих случаях, разумеется, идет о легкой снасти, имеющей одно грузило. Для дальнего заброса и заброса в ветер грузил понадобиться несколько. Дальше от крючка располагаются более тяжелые, ближе — легкие грузила. Но расстояние от нижнего грузика до крючка также варьируют в зависимости от характера поклевок. Вернемся еще раз к поплавку.
Для дальнего заброса и заброса в ветер грузил понадобиться несколько. Дальше от крючка располагаются более тяжелые, ближе — легкие грузила. Но расстояние от нижнего грузика до крючка также варьируют в зависимости от характера поклевок. Вернемся еще раз к поплавку.
Кроме «гусиного» пера при ловле карася все чаще используют поплавки самых причудливых конструкций. Некоторые из них, несомненно, заслуживают внимания.
Например, если у вас достаточно острое зрение, то можно испробовать в деле очень чуткий поплавок с антенной (рис. 4 а). Она делается из бамбука длиной 10— 12 см и толщиной около миллиметра.
Рис 4.На нее одеваются попеременно кусочки красного и белого кембрика — оплетки от радиотехнических проводов. Такая своеобразная «линейка» позволяет заметить мельчайшее притопление или всплытие поплавка.
Крепится антенна в небольшой «оливке» из пробки или окрашенного в коричневый цвет пенопласта. Оставаясь во время поклевки под водой, «оливка» сама по себе не оказывает сопротивления клюнувшей рыбе. Это обстоятельство сказывается на повышении чувствительности всей снасти. В результате при слабом клеве такой поплавок лучше других справляется со своими обязанностями.
Это обстоятельство сказывается на повышении чувствительности всей снасти. В результате при слабом клеве такой поплавок лучше других справляется со своими обязанностями.
Кстати, существует масса разновидностей описанной конструкции. Антенну заменяют пустым стержнем от авторучки, запаяв его концы и краской нанеся красно-белые полоски. Или на конец бамбуковой антенны ставят для заметности небольшой яркий шарик. Другие, ставя такой шарик, делают его передвигающимся и отмечают им границу притопления поплавка, считая, что так заметнее начало поклевки. Есть вариант, когда антенна плавно переходит в основное тело поплавка, напоминая крохотную бутылочку.
Существуют и другие конструкции. Некоторые предназначены, например, для людей с ослабленным зрением. Вот один такой поплавок (рис. 4 б, в). Он состоит из двух частей, соединенных на шарнире. Основное его тело, длиной до 8 см, толще, чем прикрепленная к его вершинке верхняя часть, которая, в свою очередь, достигает высоты в 12 см. Украинские рыболовы, придумавшие этот поплавок, красят его нижнюю часть в коричневый цвет, а верхнюю — в бело-оранжевый. При поклевке верхняя часть ложится на воду, что очень заметно.
При поклевке верхняя часть ложится на воду, что очень заметно.
Для ловли днем, в безветренную погоду, когда караси собираются в стаи и плавают у поверхности, но далеко от берега, имеется довольно оригинальный поплавок, используемый подмосковными рыболовами (рис. 4 г). Его бамбуковый шестик проходит на некотором отдалении от центра пенопластового тела и может в нем передвигаться по желанию рыболова. Нижний конец шестика огружен свинцом, а верхний при ловле находится под углом к поверхности воды. Этот угол при необходимости изменяют путем перемещения шестика.
При поднимании насадки поплавок становится вертикально, а при потяжке в сторону — ложится.
Размеры поплавка берутся по вашему желанию. Соблюдается лишь одно правило: шестик, если он вытянут вверх больше чем наполовину, должен составлять по отношению к поверхности воды угол в 45°. Регулировка поплавка достаточно нежное дело и проводится в соответствии с весом насадки.
Оценивая этот поплавок, нельзя не отметить, что ловля карася у поверхности воды пока мало популярна. Сказывается привычка и малая информированность. Но попробуйте жарким солнечным днем приглядеться к поверхности пруда, особенно там, где к воде склоняются ветви наклонившихся деревьев. Среди легкой ряби и солнечных блесток вы увидите расходящиеся круги и мелькание легких теней — это рядом с верховкой кормится и карась. И тут поплавок с наклонным шестиком будет как нельзя кстати. Лишь одно «но»: поплавок не любит порывов ветра.
Сказывается привычка и малая информированность. Но попробуйте жарким солнечным днем приглядеться к поверхности пруда, особенно там, где к воде склоняются ветви наклонившихся деревьев. Среди легкой ряби и солнечных блесток вы увидите расходящиеся круги и мелькание легких теней — это рядом с верховкой кормится и карась. И тут поплавок с наклонным шестиком будет как нельзя кстати. Лишь одно «но»: поплавок не любит порывов ветра.
Если карась держится у поверхности воды недалеко от берега, то можно просто переиначить поплавочную снасть. Для этого нужно снять грузило и попробовать ловить на медленно тонущую насадку. «Поехавший» в сторону небольшой шарообразный поплавок подскажет тем самым время подсечки.
Днем можно поймать карася и у кромки водных растений, в прогалах чистой воды, на снасть, напоминающую нахлыст. Поплавок для этого снимают, а голубоватую (желательно) леску 0,15—0,2 мм смазывают жиром. Но последние 10—15 см до крючка жиром не трогают. Этот участок лески медленно тонет, привлекая рыбу. Насадка — обычная. С воздушными насекомыми карась не особенно рад встречаться. А чтобы облегчить заброс, леску сделайте не длиннее удилища.
Насадка — обычная. С воздушными насекомыми карась не особенно рад встречаться. А чтобы облегчить заброс, леску сделайте не длиннее удилища.
Ловля на мормышку уже упоминалась. Но можно се сделать более классической, если поплавок сменить на кивок на конце удилища. Он несколько длиннее и толще, чем для зимних удочек. И делается как из упругой проволоки, так и из пружинки. На конец удилища кивки крепятся с помощью резиновой шайбы. А оканчиваются ярким шариком из пенопласта, заметным издалека. Леска пропускается обычно внутри пружинки.
Сами мормышки берутся несколько покрупнее, чем обычно используются на прудах зимой (см. раздел «Окунь»). Насадка — мотыль и опарыш. Если сохранен поплавок, то опарыш предпочтительнее — крепче держится на крючке.
С первыми похолоданиями поведение карася начинает быстро меняться. Он уходит из зоны водных растений сначала на их кромку, а затем еще в более глубокие места. Дольше мелкого осенью ловится крупный карась.
Поэтому в августе-сентябре удача больше сопутствует тому, кто, отложив поплавочные снасти, перешел на ловлю донками.
В небольших прудах наиболее уловистыми становятся глубины возле плотин, а в более крупных водоемах — ровные илистые участки на глубине, что расположились возле коряжника и затопленных русел.
Лучшей осенней насадкой, как и весной, считается навозный червь. Применяется в качестве насадки и шарик из мякоти черного хлеба, а также опарыш.
Кое-кто из ярых приверженцев поплавочной удочки ловит ею и осенью, выезжая на глубину на лодке. Для усиления клева в этом случае на снасти крепят два крючка на расстоянии 1,5—3 см друг от друга и применяют на них разную насадку. Замечено, что на нижний крючок лучше подойдет навозный червь, а на верхний шарик из теста или черного хлеба. Клюет чаще у дна, но стоит убрать верхний крючок, как вероятность поклевки нередко снижается.
Ловят карася осенью и полудонкой. Для этого поплавок передвигают почти к самой вершинке удилища, а леску по возможности удлиняют. Положив удилище на подставку, поклевку замечают по шевелению висящего в воздухе поплавка. Если же подставки нет, то поплавок пускают плавать, приблизив кончик удилища к воде.
Если же подставки нет, то поплавок пускают плавать, приблизив кончик удилища к воде.
Полудонкой можно воспользоваться и летом, во время светлых ночей. Поплавок у берега в такое время бывает заметен.
Ловля донкой возможна и без резинового амортизатора.
Существуют различные конструкции, отличающиеся оснасткой. Какая лучше? Не ошибусь, если скажу: та, которая вам понравилась, которая оказалась наиболее удобной.
Многие забыли простейшую донку. Снасть, намотанная на мотовильце небольшого колышка, сверху которого имеется проволочный шестик с дужкой, легко умещается в кармане рюкзака. Двух таких донок вполне достаточно. Леска потребуется не очень толстая: 0,2—0,25 мм, что позволит максимально снизить вес грузила и шум от заброса. При хорошем клеве это немаловажно.
Крючки также потребуются небольшие — № 5—6. На каждой донке их не больше двух, иначе неизбежны «бороды», больше времени уходит и на обслуживание снасти. Колокольчик — самодельный, из охотничьей гильзы небольшого калибра. Язычком вполне послужат гайки от детского конструктора, закрепленные при помощи лески. Ловить можно и без колокольчиков, следя за движением лески, уходящей в воду.
Язычком вполне послужат гайки от детского конструктора, закрепленные при помощи лески. Ловить можно и без колокольчиков, следя за движением лески, уходящей в воду.
Некоторые рыболовы не успокаиваются и часто совершенствуют свои донные удочки.
Применение одноручного спиннингового удилища с катушкой расширяет возможности заброса, облегчает вываживание. Вместо спиннинга можно опробовать только одну катушку, прикрепленную к колышку. Этот вариант удобен для работы с донкой, снабженной резиновым амортизатором. Чтобы поводки с крючками не захлестывались за грузило, последнее ставят на конце снасти.
Вместо грузила из свинца ставят также специальное грузило-кормушку, сделанную из конической пружины. Подобное приспособление бывает и в продаже.
При донной ловле, как и при ловле на поплавочную удочку, прикормке уделяется не последнее место. Она, как и привада, должна быть всегда свежей. И очень важно не перекормить рыбу.
А что зимой?
В любых рыболовных справочниках вы найдете утверждение, что карась зимой либо зарывается в ил, либо впадает в оцепенение, собравшись в глубоких зимовальных ямах. На это рассчитано и ведение рыбоводных хозяйств, имеющих специальные зимовальные пруды. И все же однозначный ответ я дать не решусь.
Вот уже много лет собираю сообщения о зимнем клеве карася. И география таких данных все время расширяется. Если в начале 70-х годов карась ловился со льда на Украине (может и раньше), то к середине десятилетия упоминаются зимние уловы уже в Тамбовской области, Волгоградской, затем и Московской. Казалось бы, можно сделать вывод о постепенном распространении зимнего клева карася на север и связать это со всеобщим потеплением, но, думаю, такой вывод был бы поспешным.
Из имеющихся данных видно, что не желает спать карась в достаточно больших водоемах. А большие мелиоративные пруды в средней полосе России, в том числе в Марийской республике, появились не так уж давно. Еще один повторяющийся фактор — наличие в пруду наряду с карасем окуня. Возможно, такой сильный конкурент в поисках корма и устроил карасю бессонную жизнь.
Время добавит какие-то новые аргументы, но уже можно говорить о зимней ловле карася, если соблюдены следующие условия: отсутствие заморной ситуации, недостаток летнего корма, наличие в пруду хищной рыбы, прежде всего окуня.
О клеве карася зимой на водоемах Марийской республики имеется немало сведений. Такой клев наблюдался на Шорсолинском, Ивансолинском, Регежском и Одобелякском прудах Куженерского, на Захаровском пруду Сернурского и некоторых водоемах Мари-Турекского, Советского районов. Вполне вероятно, что такие случаи имели место где-то еще.
В какую же погоду зимой карась выходит «отобедать»?
Напрасно некоторые считают, что для этого нужна оттепель. Мне не раз приходилось ловить эту рыбу и в декабре, при морозе и пронизывающем ветре. Бывает клев и в оттепели. А вот стихает он в январе-феврале.
Насадка при ловле — мотыль. Снасть — поплавочная или мормышечная удочка. Насадка должна находиться в 1—1,5 см от грунта или касаться его. В последнем случае, если ловят на мормышку без поплавка, кивок должен быть напряжен под тяжестью приманки. Поклевка — мелкое подрагивание и раскачивание кивка, затем его распрямление (поплавок всплывает). Здесь карась себе верен.
Прикормка зимой желательна, но в меру. Щепотка-другая теплой каши или пареного комбикорма — вот весь ее состав.
Щепотка-другая теплой каши или пареного комбикорма — вот весь ее состав.
Стоит ли говорить чем хорош карась на обеденном столе? Уха, жаренье и вяление — на все годна эта рыба. А знаменитые караси в сметане любимы были и царями. Между тем порадовать себя этим блюдом легко. Нужен мелкий и средний карась (чтобы прожарились и не мешали при еде косточки), обычная сковорода, смазанная маслом или любым жиром, и вылитая на рыбу незадолго до окончания процедуры жарки небольшая порция сметаны.
* Здесь и далее в подзаголовках даются марийские названия рыб.
<<<предыдущая || первая || следующая>>>
РЫБАЛКА НА «ТАЁЖКЕ», или КАРАСЬ ЗИМОЮ – БОЛЬШЕ ЧЕМ КАРАСЬ.
- Details
- Created: 14 March 2014
Зима – не помеха для заядлого рыбака, а для некоторых рыболовов только зимняя рыбалка и является настоящей. В зимнее время года ловится разная рыба, и карась – одна из этих рыб.
Хорошо карасям: не надо ходить на работу и можно спать сколько угодно, чем они и занимаются в мелких прудах и болотцах. А караси Таёжного озера, на берегу которого раскинулся популярный этно-туристический комплекс с таким же названием, никак не могут уснуть всю зиму, плавают себе подо льдом и клюют на наши мормышки – маленький кусочек лета среди сибирской зимы. Прописаны наши караси, как я уже сказал, в Таёжном озере расположенном неподалёку от Нижневартовска и Мегиона, и их здесь превеликое множество – тысячи! И если вы решили порыбачить зимой и выловить карася, то лучшего места, чем Таёжное озеро вам не найти. Что, уже не терпится разузнать подробности? Читайте дальше!
Кто-то, может, считает, что для успешной ловли нашего карася требуются какие-то особенные снасти, привады и прикормки с секретными добавками, доскональное знание мест ловли ну и богатый опыт. А на самом деле все совсем не так! Для рыбалки на Таёжном мы используем обычные удочки с простыми мормышками и надо сказать, что сама ловля не сильно отличается от любой другой вылазки на лед с мормышкой. Известно, что для зимней ловли карася основной и самой лучшей наживкой законно считается мотыль. Бытует мнение, что карась — рыба вялая и привередливая, подолгу дегустирует наживку, часто ее выплевывает. Наш карась, скорее всего, ничего об этом не слышал и жадно заглатывает приманку! А вот что касается погоды и ее воздействия на клев, то лично мне на Таёжном озере приходилось ловить его и в непроглядную метель, и в расхлябанную оттепель, и даже в ветреные морозные дни. К вечеру рюкзак заметно тяжелеет, и чувствуется усталость от сверления большого количества лунок – значит, пора домой. Сейчас, когда я пишу эти строки, где-то подо льдом Таёжного озера плавают тысячи карасей, и я точно знаю, что некоторых из них я обязательно поймаю в ближайший выходной!
Известно, что для зимней ловли карася основной и самой лучшей наживкой законно считается мотыль. Бытует мнение, что карась — рыба вялая и привередливая, подолгу дегустирует наживку, часто ее выплевывает. Наш карась, скорее всего, ничего об этом не слышал и жадно заглатывает приманку! А вот что касается погоды и ее воздействия на клев, то лично мне на Таёжном озере приходилось ловить его и в непроглядную метель, и в расхлябанную оттепель, и даже в ветреные морозные дни. К вечеру рюкзак заметно тяжелеет, и чувствуется усталость от сверления большого количества лунок – значит, пора домой. Сейчас, когда я пишу эти строки, где-то подо льдом Таёжного озера плавают тысячи карасей, и я точно знаю, что некоторых из них я обязательно поймаю в ближайший выходной!
Карась
Как поймать карася поплавочной удочкой на мелководье. Узнайте секреты рыбалки от опытного карасятника.
Секретная прикормка для пугливого карася
Самое тонкое дело при поплавочной ловле карася, правильно его закормить. Сейчас лето, карась крутится вблизи берега на мелководных участках до 1.5 метра. Поступим хитрым образом, сделаем мощный стартовый закорм, но непростой, рыбе карасю он так легко не достанется. Для этой цели применим …
Сейчас лето, карась крутится вблизи берега на мелководных участках до 1.5 метра. Поступим хитрым образом, сделаем мощный стартовый закорм, но непростой, рыбе карасю он так легко не достанется. Для этой цели применим …
Как ловить карася в мае на пруду. Едва рассвело, а компания рыболовов уже находилась в селе Урлейка Пензенской области, где была сделана остановка в надежде на поимку карася. Поговорим сегодня на тему как ловить карася в мае, о его предпочтениях, выборе снасти для данной пресноводной рыбы. Мы находимся на довольно большом пруду, в котором обитает не только карасик, но и достаточно крупный карп, белый амур, судак и даже лещ. Будем …
Узнать больше »Ловля карася на мормышку летом. 25 июня состоялась рыбалка в черте города Черняховска в Калининградской области, на речках Анграпа и Инструч. Интереснейшая и результативная ловля карася и леща проходила на мормышку маховой удочкой с боковым кивком.
Этот способ рыбалки интересен тем, что можно облавливать самые неожиданные участки водоема. Иногда рыба клюет практически вблизи самого берега, буквально в нескольких сантиметрах. …
Иногда рыба клюет практически вблизи самого берега, буквально в нескольких сантиметрах. …
Майская рыбалка в Челябинской области проходила недалеко от озера Буташ в Еткульском районе. На одном безымянном озерце рыболовы программы «Простые радости» будут ловить весеннего карася. Вода уже прогревается, у самого берега она значительно теплее, чем в метрах 3-х от линии берега, где всего лишь на десять сантиметров глубже и на пару градусов явно холоднее. На мелководья начинает выходить на нерест рыба. Хотя рыбалка …
Узнать больше »Сегодня будет затронута тема — ловля карася зимой. Эта необычная рыбалка на Урале проходила на одном из уральских водоемов, где ночью клюет карась. Карась вообщем то зимой клюет проявляя свою активность, правда не во всех прудах. В основном это водоемы, где хороший кислородный режим, имеется проточность, водится иная рыба, к примеру окунь, плотва. Преимущественно берет при сравнительно теплой с легким морозцем, ровной погоде. …
Узнать больше »АгроБиотехнологии — биотехнологии в сельском хозяйстве
Статьи и рекомендации по ловле карася
Только-только растаял снег, сошел лед с водоемов и весна потихоньку начинает вступать в свои права. Но ночью еще очень часто бывают морозы. Ранняя весна — это время начала ловли крупного весеннего карася.
Но ночью еще очень часто бывают морозы. Ранняя весна — это время начала ловли крупного весеннего карася.
Сегодня мы собрались ловить карася старым дедовским способом. Мы ловим его на фидер, который мы упростили до донки. Читать далее…
КРУПНЫЙ КАРАСЬ НА ПОПЛАВОК: СЕКРЕТЫ ЛОВЛИ
Тема сегодняшнего разговора — это ловля трофейных и исключительно трофейных карасей. В большинстве водоемов нет каких-то проблем с наличием крупного карася, но почему-то в крупных и средних озерах, как правило, рядовому рыболову попадается только мелкий и средний карасик. Есть, конечно, водоемы, где основную массу улова составляют хорошие караси, массой 300-500 грамм. Читать далее…
Среди поклонников ловли карася в прудах, только маленькая часть рыбаков предпочитает ловить этих серебристых красавцев на летние снасти с большим боковым кивком.
Чаще всего, их уловы гораздо значительнее , чем рыболовов, которые используют для ловли карася различные всевозможные донные или поплавочные снасти.
ЗА ОСЕННИМ КАРАСЕМ НА ПЛЕС
Большинство рыболовов думают, что осенью, когда вода уже стала холодной, карась прекращает клевать. Но это не совсем верно. Однозначно, караси начинают капризничать потому, что осенняя погода часто меняется.
Хотя, в центральной России довольно много водоемов, в которых карась ловится на донные снасти до ледостава в декабре. Как правило, это большие пруды с проточной водой и запруды с с большим количеством глубоких поливов. Бывает неплохой клев осенью и на реках. Но в осеннее время на реках найти места стоянок карасей очень непросто: рыба становится малоподвижной, предпочитает стоять стаей в ямках с тихим течением, и попробуй найди там этих карасей! Читать дальше…
С ФИДЕРОМ ЗА КАРАСЕМ: НАСАДКИ И ПРИКОРМКИ
В летнее время очень результативна ловля прудовых карасей с помощью фидерными снастями, особенно когда рыбак имеет хорошие навыки владения такими снастями и знает где держится карась на этом водоеме. После того, как вы выбрали водоем, где собираетесь ловить карасей, необходимо хорошо закормить то место, в которое будете затем забрасывать фидерные кормушки… Читать дальше…
После того, как вы выбрали водоем, где собираетесь ловить карасей, необходимо хорошо закормить то место, в которое будете затем забрасывать фидерные кормушки… Читать дальше…
Пруды, где водятся караси, различаются между собой по целому ряду условий, которые тем или иным образом влияют на рыбу. Есть пруды в которых проживают исключительно караси. Обычно, это водоемы с небольшими глубинами, берега которых сильно заболочены. Бывает, что такие пруды образуют целый каскад, в котором эти водоемы соединяются между собой небольшими ручейками-протоками. Зимой толщина льда достигает до самого дна, и другие рыбы, кроме карася, просто не могут выжить в подобных условиях. В таких прудах при ловле карася можно не опасаться набегов мелочи поэтому можно смело использовать любые прикормки и насадки. Однако, карась не всегда клюет на все подряд даже при отсутствии других пищевых конкурентов. Встречаются водоемы, в которых карась ловится, например, только на червяка или только на опарыша и т.
 д. Читать дальше…
д. Читать дальше…На большинстве рек, которые были когда-то запружены, развелось немалое количество серебряных карасей. Их с успехом ловят фидерной снастью вместе с другими представителями речной ихтиофауны — плотвичкой, лещом, язем, красноперкой, и другими. Но самая увлекательная и динамичная рыбалка — это ловля карасей на поплавочные удочки в окошках среди зарослей кувшинки и прочих водяных растений. Читать дальше…
КАК ПОЙМАТЬ ЗИМОЙ КАРАСЯ — ЛЕЖЕБОКУ
Большинство и написанного о зимней жизни серебряного карася не прошло проверки практикой многих рыболовов. А ведь у любителей зимней рыбалки накопился колоссальный опыт по ловле этой рыбы в зимнее время года. На многих прудах подмосковья и в начале, и в конце зимы серебряные караси клюют очень, и очень неплохо. Читать далее…
ЗИМНИЙ КАРАСЬ
С наступлением зимы иногда специально выезжаю на озера порыбачить на зимнюю удочку с поплавком. Обычно на эту снасть попадается различная мирная рыба типа плотвы, леща, сига и т.д. А не так давно начали регулярно ловиться караси — именно те караси, которые согласно всем трудам ихтиологов должны впадать в зимнюю спячку, зарываясь в илистые донные отложения, впав в состояние анабиоза. Вот о такой ловле карасей в зимнее время на зимнюю поплавочную удочку, мы и поговорим в этой статье. Читать далее…
Обычно на эту снасть попадается различная мирная рыба типа плотвы, леща, сига и т.д. А не так давно начали регулярно ловиться караси — именно те караси, которые согласно всем трудам ихтиологов должны впадать в зимнюю спячку, зарываясь в илистые донные отложения, впав в состояние анабиоза. Вот о такой ловле карасей в зимнее время на зимнюю поплавочную удочку, мы и поговорим в этой статье. Читать далее…
Множество рыбаков твердит о влиянии глобального потепления, из-за которого караси активно питаются в зимнее время. Только вот уже много лет подряд наши сибирские зимы «радуют» нас лютыми морозами, а серебряный карась замечательно клюет даже в самое глухое время, однако, количество пойманной рыбы сильно отличается от добычи при ловле по перволедью и перед самым ледоходом, но все равно же он клюет — и суровых сибирских морозов он не боится! Читать далее…
ЛОВЛЯ КАРАСЯ ВЕСНОЙ
Весенняя охота на карася наиболее результативна в тот период, когда карась сбивается в большие стаи перед нерестом и выходит на мелководье в прибрежную прошлогоднюю растительность в преднерестовый период. В это время еще не поднялись со дна водоросли, а мягкая тина, которая растет на прошлогоднем камыше, является основным кормом для карасей. Именно на это время и приходится знаменитый преднерестовый жор карася. Читать далее…
В это время еще не поднялись со дна водоросли, а мягкая тина, которая растет на прошлогоднем камыше, является основным кормом для карасей. Именно на это время и приходится знаменитый преднерестовый жор карася. Читать далее…
КАРАСЬ ВЕСНОЙ — КАК И ГДЕ ЛОВИТЬ?
Весной, когда температура воды поднимется до +12 — +13°С, крупных и средних карасей можно обнаружить в зарослях тростника. Опытным путем было установлено, что температура воды возле стеблей тростника выше на несколько градусов, чем на незаросших водной растительностью местах водоема. Стебли тростника, полые внутри и могут накапливать в себе солнечное тепло. Как раз из-за этого, лед на участках с растительностью, которая выступает из подо льда, самый тонкий и опасный. Читать дальше…
ОХОТА НА ТРОФЕЙНОГО КАРАСЯ
Ловля крупного карася по принципу «где встал там и забросил» удачи, в этом деле, не принесут. Это происходит потому, что крупные караси сильно привязаны к своим постоянным кормовым местам и местам постоянного обитания, которые они редко покидают. Поэтому, какие бы привлекательные насадки вы не забрасывали вдали от мест «постоянной дислокации» крупных карасей, все ваши попытки обречены на провал. Читать далее…
Поэтому, какие бы привлекательные насадки вы не забрасывали вдали от мест «постоянной дислокации» крупных карасей, все ваши попытки обречены на провал. Читать далее…
ЛОВЛЯ КАРАСЯ НА ПЕНОПЛАСТ
Использование пенопластовых шариков, в качестве насадки для ловли карася, это очень добычливый способ ловли этой рыбы, но, к сожалению, мало кто ловит таким способом. Такая ловля, с применением пенопласта, очень схожа с фидерной ловлей, но имеет и свои особенности, о них и будет рассказано в настоящей статье. Читать далее…
ЛОВЛЯ КАРАСЯ МАХОВОЙ УДОЧКОЙ
В настоящей статье мы поговорим о ловле карася с помощью маховой поплавочной удочки.
Удилище для карасиной ловли должно отвечать определенным критериям — она должна быть, по возможности, максимально легкой и быть довольно жесткого строя, чтобы не позволить попавшемуся на крючок карасю при первых, самых сильных рывках, забиться в прибрежные водоросли. Удилища должны быть довольно длинными — 5-6 метров, ведь трофейные караси очень осторожны и стараются держаться от рыбака, сидящего на берегу, подальше. Читать далее…
ЛОВЛЯ КАРАСЯ НА ФИДЕР
По мнению многих рыбаков, самой результативной снастью при ловле крупных карасей стали донные фидерные удочки. Эти снасти сейчас приобрели большую известность и распространение. В этой статье мы остановимся на различных вариантах ее оснастки и тактике ловли. Читать далее…
ЛОВЛЯ КАРАСЯ ПОЗДНЕЙ ОСЕНЬЮ
Существует распространенное мнение, что с поздней осени и до весны, караси впадают, в так называемую зимнюю спячку. Стаи карасей скапливаются в определенных подходящих для них местах водоема и практически прекращают питание. Однако, есть большое количество прудов и озер, в которых есть в избытке корм и кислород, что позволяет карасю зимой вести активный образ жизни. Но если в таких прудах и озерах карась активен зимой, то и поздней осенью он, в принципе, должен клевать. Читать далее…
Читать далее…
ПРИКОРМКА ДЛЯ КАРАСЯ СВОИМИ РУКАМИ
Прикормочная смесь, для привлечения карасей к месту лова, должна быть, по возможности, более естественной – обладать знакомым вкусом и запахом. Не рекомендуется добавлять в состав прикормки какие-нибудь необычные компоненты, в том числе, имеющие резкий и непривычный запах. Такие прикормочные смеси настораживают или отпугивают крупных карасей. Читать далее…
Центральные районы нашей страны богаты разной рыбой, а особенно той, которая предпочитает теплую воду. Сегодня мы будем вести рассказ о симпатичном карасином племени в озерах Воронежской области. Читать дальше…
Для многих из нас карась – рыба из рыболовного детства. Кто-то и сейчас регулярно ездит на карася, а кто-то предпочитает более предсказуемую плотву и леща, но все равно в глубине души, я уверен, совсем не прочь попасть в карасевое эльдорадо…Читать далее.
 ..
..
КАРАСЬ В НАЧАЛЕ ОСЕНИ
С приходом осени начинает изменяться список насадок, на которые реагирует осенний карась. Также, меняются места, на которых предпочитает стоять карась. Периодичность выхода на кормежку у карасей тоже изменяется. Клев карася становится непредсказуем… Читать далее…
Глупая невестка требует развода
Давным-давно сын помещика был глуп, но женился на умной и красивой невестке. Невестка чувствовала себя обиженной и требовала развода. Хозяин очень беспокоился, поэтому он пригласил джентльмена из частной школы, чтобы научить его сына некоторым знаниям. Мистер Частная школа вывел дурака из дома. Двое увидели дятла на дереве, лязгающего о дерево, и дурак спросил, что делает птица? Муж сказал: «Дятлы, дятлы, нельзя есть, когда клюешь в дверь.Идиот вспомнил эту фразу в своем сердце. Через некоторое время они увидели мертвую рыбу на земле, а вокруг клевало много птиц. В этот момент с неба спикировал орел, и птицы все улетели. Муж сказал: «Это называется приземление птицы и вставание птицы». Они прошли мимо рыбного пруда, и какие-то люди ловили рыбу в пруду. Глупый сказал: «Сэр, пойдем ловить рыбу». Муж сказал: «Сачка под рукой нет, так что к пруду не спустишься.Он встряхнул свое тело, а подойдя, вздохнул и сказал: «Шуанцяо лучше, но по дереву тяжело ходить». «Это называется бой желтохвостых карасей. Вода, ты только посмотри на людей». Пройдя некоторое время, он увидел большого буйвола, катающегося по грязи. Муж сказал: «Старая охотничья корова катится всеми четырьмя ногами к небу». берегу реки я увидел женщину, которая держала его, бамбуковая корзина ловила креветок у реки.Муж сказал: «Бамбуковые корзины рыщут по креветкам, и ни одна из них не может убежать». Когда они расстались вечером вдвоем, муж сказал дурочке и сказал: «Прощай сегодня. Везде».
Через некоторое время они увидели мертвую рыбу на земле, а вокруг клевало много птиц. В этот момент с неба спикировал орел, и птицы все улетели. Муж сказал: «Это называется приземление птицы и вставание птицы». Они прошли мимо рыбного пруда, и какие-то люди ловили рыбу в пруду. Глупый сказал: «Сэр, пойдем ловить рыбу». Муж сказал: «Сачка под рукой нет, так что к пруду не спустишься.Он встряхнул свое тело, а подойдя, вздохнул и сказал: «Шуанцяо лучше, но по дереву тяжело ходить». «Это называется бой желтохвостых карасей. Вода, ты только посмотри на людей». Пройдя некоторое время, он увидел большого буйвола, катающегося по грязи. Муж сказал: «Старая охотничья корова катится всеми четырьмя ногами к небу». берегу реки я увидел женщину, которая держала его, бамбуковая корзина ловила креветок у реки.Муж сказал: «Бамбуковые корзины рыщут по креветкам, и ни одна из них не может убежать». Когда они расстались вечером вдвоем, муж сказал дурочке и сказал: «Прощай сегодня. Везде».
Хотя этот придурок очень глупый, но у него хорошая память, поэтому он вспомнил, чему муж его учил прямо сейчас. На второй день свекор и свекровь принесли большую группу Родственники к помещику в дом для обсуждения развода.Глупая была в комнате и услышала, как кто-то стучит в дверь, и тут же громко сказала: «Дятел, дятел, клюют, отворяют дверь и есть не могут.Группа людей снаружи слушала это, и они думали: Этот человек не глупый, кажется, немного знающий. Все вошли в дом и все заняли свои места. В этот момент снаружи вошел хозяин, и все в доме встали, чтобы поприветствовать его. Глупец громко сказал: «Это называется птичка приземляется и птичка встает». Все были потрясены, и хвалили его за ловкость и сообразительность. его, и тут же приказал кухне готовить и угощать гостей.На какое-то время еда была расставлена на столе, прежде чем были расставлены миски и палочки для еды, хозяин попросил кого-нибудь поставить миски и палочки для еды. В это время глупый мальчик снова сказал: «На твоей руке нет сети, ты не можешь спуститься к пруду». В это время ни у кого не было сомнений. Оказывается, этот пацан раньше притворялся дураком, а на самом деле он, можно сказать, хороший человек.
На второй день свекор и свекровь принесли большую группу Родственники к помещику в дом для обсуждения развода.Глупая была в комнате и услышала, как кто-то стучит в дверь, и тут же громко сказала: «Дятел, дятел, клюют, отворяют дверь и есть не могут.Группа людей снаружи слушала это, и они думали: Этот человек не глупый, кажется, немного знающий. Все вошли в дом и все заняли свои места. В этот момент снаружи вошел хозяин, и все в доме встали, чтобы поприветствовать его. Глупец громко сказал: «Это называется птичка приземляется и птичка встает». Все были потрясены, и хвалили его за ловкость и сообразительность. его, и тут же приказал кухне готовить и угощать гостей.На какое-то время еда была расставлена на столе, прежде чем были расставлены миски и палочки для еды, хозяин попросил кого-нибудь поставить миски и палочки для еды. В это время глупый мальчик снова сказал: «На твоей руке нет сети, ты не можешь спуститься к пруду». В это время ни у кого не было сомнений. Оказывается, этот пацан раньше притворялся дураком, а на самом деле он, можно сказать, хороший человек. Слуга положил перед тестем палочки для еды. Дурак увидел его, вздохнул и сказал: «Двойной мост лучше, по одинарному дереву ходить тяжело.»
Слуга положил перед тестем палочки для еды. Дурак увидел его, вздохнул и сказал: «Двойной мост лучше, по одинарному дереву ходить тяжело.»
выпить. Допустим, что глупая невестка не знала, как идет развод, поэтому подошла и заглянула украдкой в щель двери. Глупый увидел это и сказал жене: «Желтохвостый карась борется с водой, только глядя на людей.» Услышав это, тесть Свекровь и все приглашенные родственники сильно покраснели, думая, что это малыш такой могущественный, и издевался над своей женой и ее девственной семьей и обвинял их в том, что они хуже.Все впечатлились глуповатым талантом, долго стеснялись оставаться, торопливо поели и ушли. Старый тесть поскользнулся и упал во двор. Глупый громко запел: «Старая охотничья корова катится с поднятыми ногами». Гости устыдились, помогли старому тестю и угрюмо ушли. Глупые погнались и закричали: «Бамбуковые палочки рыщут в поисках креветок. Не могу бежать». Гости так испугались, что души их разлетелись, а глупые закричали: «Я еще ни слова не сказал. : «Прощай сегодня, встречи негде».
: «Прощай сегодня, встречи негде».
Множество путей, ведущих к системе NOS/NO в сердце рыбы
Abstract
К костистым рыбам часто относятся с интересом из-за замечательной способности некоторых видов переносить даже драматические события. стрессы, как внутренние, так и внешние, как в случае колебаний наличия O 2 и температурных режимов.Эти события естественным образом переживают многие виды рыб в различных временных масштабах, но теперь они усугубляются растущими изменениями окружающей среды.Это еще больше бросает вызов внутренней способности животных справляться со стрессом. Сердце имеет решающее значение для реакции на стресс, поскольку правильная модуляция сердечной функции обеспечивает перфузию крови во всем организме, особенно в органах дыхания и головном мозге. В сердечных клетках активируются ключевые сигнальные пути для поддержания молекулярного равновесия, что улучшает стрессоустойчивость. У рыб система синтазы оксида азота (NOS)/оксида азота (NO) имеет основополагающее значение для модуляции основных функций сердца и участвует в контроле многих адаптивных реакций на стресс, в том числе связанных с вариациями O 2 и термическими изменениями. режимы.В этом обзоре мы стремимся проиллюстрировать, объединяя классическую и новую литературу, современные знания о системе NOS/NO как важнейшем компоненте сердечных молекулярных механизмов, поддерживающих стрессоустойчивость и адаптацию, тем самым предоставляя некоторые виды, такие как толерантные карповые, с высокой стрессоустойчивостью.
режимы.В этом обзоре мы стремимся проиллюстрировать, объединяя классическую и новую литературу, современные знания о системе NOS/NO как важнейшем компоненте сердечных молекулярных механизмов, поддерживающих стрессоустойчивость и адаптацию, тем самым предоставляя некоторые виды, такие как толерантные карповые, с высокой стрессоустойчивостью.
Ключевые слова: нитрергическая система, сердце рыб, работа сердца, многократный стресс, гипоксия, температурные изменения, АМПК
1. Введение
Живые организмы постоянно подвергаются стрессу.После определения Сейлем стресса как «неспецифической реакции организма на любое предъявляемое к нему требование» и последующего отклонения различной степени реакции от эустресса к дистрессу [1,2] десятилетия исследований установили, что способность преодоление стрессозависимых проблем представляет собой основной механизм поддержания гомеостаза организма. Многие факторы формируют реакцию на стресс, что делает сценарий очень сложным. В попытке нарисовать структуру, в которой резюмируются сети и цепи, участвующие в перекрестных помехах между окружающей средой и организмами, было разработано несколько связанных со стрессом концепций, таких как стрессом (каталог генов и их продуктов, участвующих в неадаптивных реакциях). стресс-реакция) и стрессотоп (адаптационный фон, включающий цепи молекулярных медиаторов, участвующих в стресс-реакции, вплоть до популяционного уровня) [3].Важным аспектом в контексте адаптивного ответа является то, что множественные стрессы часто сходятся, бросая вызов организму, и это вызывает реакцию, которая является результатом многих событий, происходящих на многоуровневом уровне, от всего организма до молекулярной сигнализации [3]. . Это случай родственных стрессов, таких как гипоксия и температура, которые, особенно в водной среде, действуют одновременно. Изменения температуры обратно пропорционально влияют на растворимость в воде O 2 ; в то же время термические модификации влияют на потребность в энергии водных экзотермических видов.
В попытке нарисовать структуру, в которой резюмируются сети и цепи, участвующие в перекрестных помехах между окружающей средой и организмами, было разработано несколько связанных со стрессом концепций, таких как стрессом (каталог генов и их продуктов, участвующих в неадаптивных реакциях). стресс-реакция) и стрессотоп (адаптационный фон, включающий цепи молекулярных медиаторов, участвующих в стресс-реакции, вплоть до популяционного уровня) [3].Важным аспектом в контексте адаптивного ответа является то, что множественные стрессы часто сходятся, бросая вызов организму, и это вызывает реакцию, которая является результатом многих событий, происходящих на многоуровневом уровне, от всего организма до молекулярной сигнализации [3]. . Это случай родственных стрессов, таких как гипоксия и температура, которые, особенно в водной среде, действуют одновременно. Изменения температуры обратно пропорционально влияют на растворимость в воде O 2 ; в то же время термические модификации влияют на потребность в энергии водных экзотермических видов. И O 2 , и температура динамически изменяются в течение дней, сезонов, лет и даже в географическом масштабе для мигрирующих видов, так что животные постоянно и физиологически подвергаются потребности адаптации [4,5]. В последнее время стресс, вызванный изменениями в воде O 2 и температуре, резко возрастает из-за интенсивных манипуляций человека с окружающей средой и добавления других факторов, например, неестественных химических веществ (см. для справки [6 ]).Это еще больше бросает вызов внутренней толерантности животного.
И O 2 , и температура динамически изменяются в течение дней, сезонов, лет и даже в географическом масштабе для мигрирующих видов, так что животные постоянно и физиологически подвергаются потребности адаптации [4,5]. В последнее время стресс, вызванный изменениями в воде O 2 и температуре, резко возрастает из-за интенсивных манипуляций человека с окружающей средой и добавления других факторов, например, неестественных химических веществ (см. для справки [6 ]).Это еще больше бросает вызов внутренней толерантности животного.
При стрессовых воздействиях, одиночных или множественных, выживание организма обеспечивается адаптивной гибкостью важнейших органов, таких как мозг и сердце, основные функции которых должны строго сохраняться. Соответственно, эти органы с интересом рассматриваются как биологические тесты в исследованиях, связанных с адаптацией к стрессу.
Большое количество литературы описывает механизмы, которые у водных видов поддерживают адаптацию сердца к условиям окружающей среды, уделяя внимание не только верхним и нижним пределам этой адаптации, но и молекулярным путям, которые задействуются во время воздействия к многочисленным вызовам, событие, которое становится все более и более частым в поврежденной природной среде. Недавним примером является сердечный транскриптомный ответ, описанный у личинок Fundulus grandis , развивающихся на четыре комбинированных стрессора (доступность O 2 , температура, соленость и полициклические ароматические углеводороды) [7]. Было обнаружено, что каждый отдельный стресс, применяемый отдельно, влияет на сердце с точки зрения гемодинамики и развития от удара к удару. В то же время сочетание стресса потенцирует как положительное, так и отрицательное воздействие на сердце, воздействуя на канонические пути, участвующие в сократительной способности сердца, вазомоторике и пролиферации кардиомиоцитов.Эти пути включают сердечную нитрергическую систему [7]. Как показано во многих работах, в сердце эта система находится на перекрестке многих чувствительных к стрессу цепей [8,9,10,11,12,13]. Он находится под модуляцией, вызванной стрессом, и в то же время имеет решающее значение для формирования реакции на стресс, как адаптивной, так и неадекватной.
Недавним примером является сердечный транскриптомный ответ, описанный у личинок Fundulus grandis , развивающихся на четыре комбинированных стрессора (доступность O 2 , температура, соленость и полициклические ароматические углеводороды) [7]. Было обнаружено, что каждый отдельный стресс, применяемый отдельно, влияет на сердце с точки зрения гемодинамики и развития от удара к удару. В то же время сочетание стресса потенцирует как положительное, так и отрицательное воздействие на сердце, воздействуя на канонические пути, участвующие в сократительной способности сердца, вазомоторике и пролиферации кардиомиоцитов.Эти пути включают сердечную нитрергическую систему [7]. Как показано во многих работах, в сердце эта система находится на перекрестке многих чувствительных к стрессу цепей [8,9,10,11,12,13]. Он находится под модуляцией, вызванной стрессом, и в то же время имеет решающее значение для формирования реакции на стресс, как адаптивной, так и неадекватной.
В следующих параграфах основное внимание уделяется физиологическим реакциям сердца рыб на ограничения O 2 и колебаниям температуры, которые используются в качестве примеров большого разнообразия проблем, с которыми животные должны сталкиваться естественным образом. В этом контексте система синтазы оксида азота (NOS)/оксид азота (NO) будет обсуждаться как важнейший компонент молекулярных механизмов, формирующих реакцию сердца рыб на стресс, особенно у видов, характеризующихся высокой адаптивной гибкостью, т.к. в случае карповых (). Предварительное описание системы NOS/NO в контроле основной сердечной функции и важная информация о сердечной реакции на O 2 и изменениях температуры у костистых рыб будут также предоставлены для незнакомых читателей, так как эта информация является отправной точкой для все оценки касались участия нитрергической системы в реакции на стресс.
В этом контексте система синтазы оксида азота (NOS)/оксид азота (NO) будет обсуждаться как важнейший компонент молекулярных механизмов, формирующих реакцию сердца рыб на стресс, особенно у видов, характеризующихся высокой адаптивной гибкостью, т.к. в случае карповых (). Предварительное описание системы NOS/NO в контроле основной сердечной функции и важная информация о сердечной реакции на O 2 и изменениях температуры у костистых рыб будут также предоставлены для незнакомых читателей, так как эта информация является отправной точкой для все оценки касались участия нитрергической системы в реакции на стресс.
Графическое представление содержания обзора. В условиях ограничений O 2 и температурных колебаний система синтазы оксида азота (NOS)/оксид азота (NO), прямо или косвенно взаимодействуя с критическими киназами, например, AMP-активируемой протеинкиназой (AMPK), модулирует молекулярные механизмы. что в сердце рыбы поддерживают стрессоустойчивость и адаптацию.
2. Система NOS/NO и сердце рыбы
NO является газотрансмиттером, генерируемым семейством изоферментов NOS, которое включает конститутивную эндотелиальную (eNOS) и нейрональную (nNOS) и индуцибельную (iNOS) изоформы.Используя молекулярный O 2 и NADPH в качестве незаменимых кофакторов, эти ферменты превращают L-аргинин в L-цитруллин и NO.
NO оказывает свое физиологическое действие либо посредством механизмов, зависящих от растворимой гуанилатциклазы (рГЦ), либо реагируя с гемами, тиолами или аминами, образуя нитрозил железа (FeNO), S-нитрозо (SNO) и N-нитрозо (NNO) соединений [14]. NO имеет очень короткий период полураспада. Он быстро метаболизируется до нитрита в реакции с O 2 [15] и инактивируется путем окисления до нитрата в реакции с оксигенированным гемоглобином (Hb) и миоглобином (Mb).В гипоксических и/или кислых условиях нитрит может представлять собой резервуар NO, поскольку он может быть восстановлен обратно до NO различными неферментативными и ферментативными путями. К ним относятся кислотное диспропорционирование и реакция с эндогенными нитритредуктазами, такими как дезоксигенированный Hb, Mb, нейроглобин, цитоглобин, ксантиноксидоредуктаза, eNOS и некоторые митохондриальные ферменты [16].
К ним относятся кислотное диспропорционирование и реакция с эндогенными нитритредуктазами, такими как дезоксигенированный Hb, Mb, нейроглобин, цитоглобин, ксантиноксидоредуктаза, eNOS и некоторые митохондриальные ферменты [16].
Сердце является основным производителем NO и одновременно мишенью его действия. Генерируемый миокардиальными и немиокардиальными тканями, NO вызывает аутокринно-паракринный контроль сердечных ответов от удара к удару, краткосрочных и долгосрочных [17,18].
Хотя в литературе можно найти неоднозначные результаты, в настоящее время признано, что, как и у млекопитающих, в сердце рыб присутствует функциональная система NOS/NO [19,20,21,22]. nNOS и iNOS были идентифицированы у костистых рыб. Более того, физио-фармакологические и иммунодетектирующие подходы указывают на присутствие «eNOS-подобной» изоформы в сердце нескольких видов. Наличие канонической eNOS у рыб остается под вопросом. Однако Andreakis и соавт. [23] предположили, что изоформа nNOS с эндотелиально-подобным консенсусом может охватывать некоторые функциональные особенности, типичные для изоформы eNOS. В качестве примера предполагается, что консенсусная последовательность миристоилирования в nNOS миноги и в iNOS рыбок данио выполняет функции, сравнимые с eNOS млекопитающих [24].
В качестве примера предполагается, что консенсусная последовательность миристоилирования в nNOS миноги и в iNOS рыбок данио выполняет функции, сравнимые с eNOS млекопитающих [24].
Сердечные БДУ были зарегистрированы у эвритермных видов рыб ( Thunnus thynnus thynnus [25]; Anguilla anguilla ( A. anguilla ) [25,26]; Carassius auratus () () 11]), адаптированные к холоду антарктические костистые рыбы (ледяная рыба Chaenocephalus aceratus ( C.aceratus ), гемоглобина Chionodraco hamatus ( C. hamatus ), краснокровных Trematomus bernacchii ( T. bernacchii ) [27,28]), и двоякодышащих рыб ( Pro) Protopterus annectens [10]). У этих видов фермент в основном экспрессируется в эндотелии эндокарда (ЭЭ), что указывает на то, что эта ткань является важным паракринным источником NO. Он также присутствует, но в меньшей степени, в миокардиоцитах, которые могут действовать как дополнительный аутокринный источник NO. Интересно, что NOS также обнаруживаются в желудочковом висцеральном перикарде (т.е. в эпикарде), что свидетельствует о том, что эта ткань является источником нитрергической передачи сигналов. Эта экстенсивная внутрисердечная экспрессия изоформ NOS считается показателем способности NO взаимодействовать с рядом клеточных мишеней для достижения пространственно-временной модуляции сердечной функции в краткосрочных и долгосрочных ответах, в исходных условиях и в наличие физической и химической стимуляции [21,30].
Интересно, что NOS также обнаруживаются в желудочковом висцеральном перикарде (т.е. в эпикарде), что свидетельствует о том, что эта ткань является источником нитрергической передачи сигналов. Эта экстенсивная внутрисердечная экспрессия изоформ NOS считается показателем способности NO взаимодействовать с рядом клеточных мишеней для достижения пространственно-временной модуляции сердечной функции в краткосрочных и долгосрочных ответах, в исходных условиях и в наличие физической и химической стимуляции [21,30].
Базальный нитрергический контроль был зарегистрирован в сердце различных видов рыб. У стальноголовой форели ( Oncorhynchus mykiss ( O. mykiss )) введение экзогенного NO увеличивает скорость расслабления миокарда, что, в свою очередь, влияет на продолжительность изометрических сокращений и мышечную сократимость. Этот NO-зависимый эффект связан с частотой сокращений, так как продолжительность сокращений уменьшается на 25% при частоте 20 ударов в минуту (уд/мин), но только на 5% при 80 ударах в минуту [31]. У угря [32], лосося [33] и золотой рыбки [11] тоническое высвобождение аутокринного NO негативно модулирует основные механические характеристики сердца. Кроме того, NO влияет на механизм Франка-Старлинга (лосось [33]; угорь [34]; золотая рыбка [11]), фундаментальный признак сердца, который у всех позвоночных позволяет миокарду увеличивать ударный объем (УО) и, как следствие, сердечный выброс (СО) в ответ на увеличение венозного возврата (преднагрузка). Как было обнаружено в изолированном и перфузированном сердце золотых рыбок, чувствительность к давлению наполнения снижается, когда образование NO предотвращается ингибированием NOS через L-NMMA или, ниже по течению, когда продукция цГМФ блокируется ODQ [11], таким образом вовлекая классический NO/ цГМФ-зависимый каскад.Как и в недавних данных Carnevale et al. [31] у стальноголовой форели, у угря нитрергический контроль реакции Франка-Старлинга связан с усилением релаксации, возможно, из-за обратного захвата кальция насосами SERCA2a, контролируемыми S-нитрозилированием фосфоламбана.
У угря [32], лосося [33] и золотой рыбки [11] тоническое высвобождение аутокринного NO негативно модулирует основные механические характеристики сердца. Кроме того, NO влияет на механизм Франка-Старлинга (лосось [33]; угорь [34]; золотая рыбка [11]), фундаментальный признак сердца, который у всех позвоночных позволяет миокарду увеличивать ударный объем (УО) и, как следствие, сердечный выброс (СО) в ответ на увеличение венозного возврата (преднагрузка). Как было обнаружено в изолированном и перфузированном сердце золотых рыбок, чувствительность к давлению наполнения снижается, когда образование NO предотвращается ингибированием NOS через L-NMMA или, ниже по течению, когда продукция цГМФ блокируется ODQ [11], таким образом вовлекая классический NO/ цГМФ-зависимый каскад.Как и в недавних данных Carnevale et al. [31] у стальноголовой форели, у угря нитрергический контроль реакции Франка-Старлинга связан с усилением релаксации, возможно, из-за обратного захвата кальция насосами SERCA2a, контролируемыми S-нитрозилированием фосфоламбана. [28].
[28].
Кроме того, в сердце больных рыб NO влияет на реакцию миокарда на растяжение. Это наблюдалось у атлантического лосося ( Salmo salar ( S. salar )) при ИСА (инфекционной лососевой анемии), при которой сердце характеризуется нарушением работы, коррелирующим с тяжестью патологии.У больных рыб функциональное повреждение сердца обратимо, когда доступность NO для тканей снижается за счет селективного ингибирования iNOS. Напротив, у здоровых животных при лишении NO не наблюдается никаких эффектов. Это позволяет предположить, что при наличии инфекций NO, образующийся в результате индукции iNOS, может быть механистически связан с хрупкостью сердца вследствие заболевания [33].
Мембранная проницаемость NO позволяет ему свободно диффундировать и контролировать соседние клетки. В бессосудистом или плохо васкуляризированном сердце рыб ЭЭ представляет собой основной источник NO.Данные нашей лаборатории показывают, что этот NO, полученный из ЭЭ, является обязательным для сократительных эффектов, вызываемых многими эндолюминальными химическими стимулами, такими как ацетилхолин, ангиотензин II, вазостатин-1 и β3-адренорецептор [32,35,36,37].
Следует отметить, что NO, вырабатываемый в сердце, может опосредовать действия, удаленные от места образования, благодаря хранению в крови в виде нитритов. У рыб, по сравнению с наземными животными, пул нитритов может быть обогащен за счет поглощения нитритов из окружающей среды через дыхательные поверхности [16].Кроме того, как наблюдается у карася ( Carassius carassius ( C. carassius )), пул нитритов в тканях может быть дополнительно увеличен за счет активности нитратредуктазы, обычно опосредованной ксантиноксидоредуктазой и Mb [38]. Это приводит к повышенной доступности NO с последующим большим влиянием на сердечно-сосудистую функцию. Этот механизм имеет особое значение при глубокой гипоксии и аноксии, когда, будучи NOS-ферментами, неспособными продуцировать NO из-за отсутствия O 2 , он способствует кардиопротекции [38,39].Данные по рыбкам данио подтверждают эту возможность, показывая, что воздействие воды № 2 сопровождается значительным образованием NO вместе с расширением сосудов и снижением артериального давления [40]. В присутствии гипоксии окружающей среды эти явления могут усиливаться за счет стимуляции восстановления нитритов до NO (см. ссылки в [11]).
В присутствии гипоксии окружающей среды эти явления могут усиливаться за счет стимуляции восстановления нитритов до NO (см. ссылки в [11]).
Роль NO 2 в сердечном нитрергическом контроле была изучена Cerra и коллегами у угря A. anguilla и у ледяной рыбы C.hamatus , два вида, характеризующиеся крайне разными экофизиологическими признаками [41]. Угорь является эвритермным и эвригалинным костистым организмом, который испытывает значительные колебания в окружающей среде O 2 . Напротив, ледяная рыба является экстремальным стенотермом, эндемичным для стабильных, ледяных и богатых кислородом антарктических вод. У угря нитрит отрицательно влияет на сократительную способность изолированного и перфузируемого сердца. Это происходит путем модулирования активности NOS и пути цГМФ/ПКГ.Более того, нитрит влияет на реакцию Франка-Старлинга посредством механизма, который задействует путь NO/cGMP/PKG и требует S-нитрозилирования белка [42]. В отличие от угря, у ледяной рыбы нитрит вызывает NOS-зависимое увеличение сократительной способности, подобное эффекту, вызываемому NO. У ледяной рыбы отсутствует Hb, ключевой белок гомеостаза NO, который способен не только очищать NO, но и генерировать его из NO 2 (ссылки см. в [41]). Было высказано предположение, что у ледяной рыбы восстановление NO 2 до NO происходит с помощью сердечного Mb, который у этой рыбы может представлять преобладающую форму редуктазы NO 2 [41].
У ледяной рыбы отсутствует Hb, ключевой белок гомеостаза NO, который способен не только очищать NO, но и генерировать его из NO 2 (ссылки см. в [41]). Было высказано предположение, что у ледяной рыбы восстановление NO 2 до NO происходит с помощью сердечного Mb, который у этой рыбы может представлять преобладающую форму редуктазы NO 2 [41].
2.1. Гипоксия
Водная гипоксия (уровни O 2 : ≤2,8 мг L -1 ; [43]) является основным ограничивающим фактором окружающей среды. Водные животные регулярно испытывают его либо хронически, либо на ежедневной или сезонной основе, и являются результатом сложных процессов, включая смешивание, воздухо-водяной обмен и колебания в модели производства и потребления O 2 [44,45]. Гипоксия распространена в районах, характеризующихся низким перемешиванием или ограничением света (например, болота с густой растительностью, затопленные леса и глубокие воды), а также в тропических пресных водах, где высокие температуры усиливают разложение органических веществ, снижая тем самым растворимость O 2 [46]. ].В последние годы водная гипоксия увеличивается из-за антропогенных воздействий и изменений климата [47,48], с тяжелым воздействием на отдельные организмы, сообщества и экосистемы. Наличие специализированных бранхиальных нейроэпителиальных клеток (НЭК) позволяет костистым «ощущать» кислородные изменения в водной/кровяной среде [49]. Рыбы демонстрируют очень широкий спектр чувствительности к O 2 , переходя от видов, резко страдающих от лишения O 2 , к видам, способным переносить гипоксическую или даже бескислородную среду.Например, лосось и тунец в значительной степени полагаются на аэробный метаболизм для быстрого и продолжительного плавания, будучи чрезвычайно чувствительными к гипоксии [50, 51], в то время как угорь и миксина хорошо проявляют себя также при низких уровнях O 2 [52, 53]. Представители карповых, такие как золотая рыбка ( C. auratus ) и карась ( C. carassius ), являются одними из наиболее устойчивых к гипоксии позвоночных, способных пережить сокращение или даже лишение в течение длительных периодов времени.
].В последние годы водная гипоксия увеличивается из-за антропогенных воздействий и изменений климата [47,48], с тяжелым воздействием на отдельные организмы, сообщества и экосистемы. Наличие специализированных бранхиальных нейроэпителиальных клеток (НЭК) позволяет костистым «ощущать» кислородные изменения в водной/кровяной среде [49]. Рыбы демонстрируют очень широкий спектр чувствительности к O 2 , переходя от видов, резко страдающих от лишения O 2 , к видам, способным переносить гипоксическую или даже бескислородную среду.Например, лосось и тунец в значительной степени полагаются на аэробный метаболизм для быстрого и продолжительного плавания, будучи чрезвычайно чувствительными к гипоксии [50, 51], в то время как угорь и миксина хорошо проявляют себя также при низких уровнях O 2 [52, 53]. Представители карповых, такие как золотая рыбка ( C. auratus ) и карась ( C. carassius ), являются одними из наиболее устойчивых к гипоксии позвоночных, способных пережить сокращение или даже лишение в течение длительных периодов времени. 54]. Разнообразие специализаций способствует борьбе с гипоксией; к ним относятся поведенческие, физиологические, биохимические и молекулярные реакции, которые позволяют либо увеличить поглощение O 2 из окружающей среды, либо ограничить потенциально опасные последствия для клеток, тканей и органов [54,55].
54]. Разнообразие специализаций способствует борьбе с гипоксией; к ним относятся поведенческие, физиологические, биохимические и молекулярные реакции, которые позволяют либо увеличить поглощение O 2 из окружающей среды, либо ограничить потенциально опасные последствия для клеток, тканей и органов [54,55].
В сердце общей реакцией на гипоксию является брадикардия вместе с угнетением сократительной способности миокарда и скорости потребления O 2 . Эти эффекты особенно очевидны у видов, не переносящих гипоксию [56,57,58]. Напротив, устойчивые к гипоксии виды, такие как карась и золотая рыбка, сохраняют нормальную сердечную функцию и даже усиливают ее, активируя сложный молекулярный механизм, который известен лишь частично [11, 59, 60]. Как видно на ex vivo изолированных и работающих препаратах сердца золотых рыбок, воздействие острой гипоксии сопровождается повышенной чувствительностью миокарда к увеличению преднагрузки (т.э., механизм Франка–Старлинга) [11].
Гемодинамические изменения, возникающие в условиях низкого содержания кислорода, считаются решающими для поддержания функционального и метаболического взаимодействия между органами и тканями, обеспечивающего толерантность к гипоксии всего организма [11]. При воздействии низких уровней O 2 золотые рыбки полагаются на большие запасы гликогена в печени и мышцах, снижают метаболизм и избегают молочнокислого ацидоза путем преобразования лактата в этанол и CO 2 , который быстро выводится через жабры [54,61, 62].Сохраненная или даже повышенная сердечно-сосудистая функция не только способствует мобилизации глюкозы из запасов гликогена в печени, но и позволяет транспортировать лактат в мышцы для его превращения в этанол [59].
O 2 доставка к сердцу обязательна для поддержания работы сердца [63,64]. У рыб присутствуют различные типы кровоснабжения и сердечной архитектуры. В зависимости от вида желудочек может состоять исключительно из губчатого миокарда, перфузируемого малооксигенированной венозной лакунарной кровью, или может включать наружный слой компактной ткани различной толщины, перфузируемый O 2 богатой кровью из коронарное кровообращение (ссылки и подробности о структурных характеристиках сердца рыб см. в [65,66,67]).В венозной крови, достигающей spongiosa , сатурация O 2 может снижаться примерно с 25% при нормоксии до примерно 3% при гипоксии [68,69,70]. Таким образом, губчатый миокард может подвергаться воздействию очень низких значений O 2 , что требует специфической тканевой адаптации. Действительно, низкая чувствительность к гипоксии была продемонстрирована у spongiosa стальноголовой форели ( O. mykiss ) по отношению к компактному миокарду [71].
в [65,66,67]).В венозной крови, достигающей spongiosa , сатурация O 2 может снижаться примерно с 25% при нормоксии до примерно 3% при гипоксии [68,69,70]. Таким образом, губчатый миокард может подвергаться воздействию очень низких значений O 2 , что требует специфической тканевой адаптации. Действительно, низкая чувствительность к гипоксии была продемонстрирована у spongiosa стальноголовой форели ( O. mykiss ) по отношению к компактному миокарду [71].
2.2. Система NOS/NO в сердце рыб в условиях гипоксии
Общим эффектом активации системы NOS/NO в сердце в условиях гипоксии является ограничение митохондриального потребления O 2 для конкурентного связывания NO с O 2 к CytC оксидазе [72].Когда доступность O 2 снижается, это способствует сохранению эффективности миокарда за счет увеличения силы, генерируемой на потребление O 2 [73,74].
Сравнение сердечной реакции на низкий O 2 у рыб, не переносящих гипоксию, и рыб, толерантных к гипоксии, было проведено Педерсеном и сотрудниками с использованием полосок желудочков форели и золотых рыбок [57]. У обоих видов NO, образующийся при активации NOS, подавляет частоту дыхания и способствует повышению эффективности работы миокарда.Однако, когда NO образуется в результате превращения нитрита, у этих двух видов наблюдаются разные реакции. Действительно, у форели, но не у золотых рыбок, потребление миокарда O 2 снижено без изменения развития силы. Это связано с различиями в сродстве к кислороду, а затем в нитритредуктазной способности миокардиального Mb. При меньшем количестве доступного O 2 Mb форели может легко деоксигенировать, таким образом образуя NO из нитрита, в то время как Mb золотой рыбки, оставаясь насыщенным O 2 , предотвращается за счет восстановления нитрита [57].
У обоих видов NO, образующийся при активации NOS, подавляет частоту дыхания и способствует повышению эффективности работы миокарда.Однако, когда NO образуется в результате превращения нитрита, у этих двух видов наблюдаются разные реакции. Действительно, у форели, но не у золотых рыбок, потребление миокарда O 2 снижено без изменения развития силы. Это связано с различиями в сродстве к кислороду, а затем в нитритредуктазной способности миокардиального Mb. При меньшем количестве доступного O 2 Mb форели может легко деоксигенировать, таким образом образуя NO из нитрита, в то время как Mb золотой рыбки, оставаясь насыщенным O 2 , предотвращается за счет восстановления нитрита [57].
У стальноголовой форели роль нитрергической системы в механической реакции сердца на низкий O 2 была дополнительно исследована Carnevale и коллегами [31]. При воздействии донора NO (SNP) на полоски губчатого желудочка от животных, акклиматизированных к низкому содержанию кислорода (PO 2 = 8 кПа), они обнаружили, что гипоксическая акклиматизация почти не влияет на частотно-зависимый NO-зависимый эффект на продолжительность сокращения и мышечную сократимость. Приведенные выше исследования показывают, что у форели воздействие гипоксии не оказывает существенного влияния на изометрическую сократимость сердца в ответ на NO.Однако авторы не исключают возможности того, что использование мышечных полосок может не выявить дополнительных эффектов NO, которые могут сохраняться в препарате цельного сердца, ближе к ситуации in vivo [31,57]. В соответствии с этим данные, полученные на золотых рыбках с использованием ex vivo изолированных и перфузированных препаратов рабочего сердца, метода, который предотвращает ограничения, связанные с использованием ограниченных частей органа, показывают, что потенцированная базальная активность, типичная для острого O 2 , сопровождается повышенной экспрессией NOS в миокарде [11].Возможность того, что более выраженный фермент генерирует большее количество NO и это, в свою очередь, влияет на работу миокарда, подтверждается доказательствами того, что удаление NO с помощью PTIO, а также ингибирование NOS с помощью L-NMMA снижает гипоксически-зависимое повышение сократительной способности.
Приведенные выше исследования показывают, что у форели воздействие гипоксии не оказывает существенного влияния на изометрическую сократимость сердца в ответ на NO.Однако авторы не исключают возможности того, что использование мышечных полосок может не выявить дополнительных эффектов NO, которые могут сохраняться в препарате цельного сердца, ближе к ситуации in vivo [31,57]. В соответствии с этим данные, полученные на золотых рыбках с использованием ex vivo изолированных и перфузированных препаратов рабочего сердца, метода, который предотвращает ограничения, связанные с использованием ограниченных частей органа, показывают, что потенцированная базальная активность, типичная для острого O 2 , сопровождается повышенной экспрессией NOS в миокарде [11].Возможность того, что более выраженный фермент генерирует большее количество NO и это, в свою очередь, влияет на работу миокарда, подтверждается доказательствами того, что удаление NO с помощью PTIO, а также ингибирование NOS с помощью L-NMMA снижает гипоксически-зависимое повышение сократительной способности. . Более того, в условиях гипоксии ингибирование NOS L-NMMA не изменяет реакцию Франка-Старлинга сердца золотой рыбки. Напротив, значительное снижение чувствительности миокарда к растяжению наблюдается, если NO удаляется из ткани с помощью PTIO и если рГЦ ингибируется с помощью ODQ.Это имеет значение, поскольку указывает на то, что эффекты, вызываемые NO, включают каскад цГМФ, но не зависят от NOS, что требует других путей для образования NO [75].
. Более того, в условиях гипоксии ингибирование NOS L-NMMA не изменяет реакцию Франка-Старлинга сердца золотой рыбки. Напротив, значительное снижение чувствительности миокарда к растяжению наблюдается, если NO удаляется из ткани с помощью PTIO и если рГЦ ингибируется с помощью ODQ.Это имеет значение, поскольку указывает на то, что эффекты, вызываемые NO, включают каскад цГМФ, но не зависят от NOS, что требует других путей для образования NO [75].
Молекулярные механизмы, которые позволяют NO играть роль в реакции миокарда на гипоксию, у рыб определены лишь частично. В сердце млекопитающих в исходных условиях и в присутствии низкого O 2 путь PI3-K/Akt контролирует активность eNOS и, таким образом, генерацию NO и последующую NO-зависимую передачу сигналов, защищая сердце [76, 77].То же самое происходит и у устойчивых к гипоксии рыб, у которых, как это наблюдается в перфузированном сердце золотой рыбки, воздействие низких доз O 2 сопровождается активацией этих киназ [75]. Интересно, что в гипоксическом сердце золотых рыбок молекулярные события генерации NO ниже по течению исключают участие цГМФ-зависимой передачи сигналов [75]. Пути, не зависящие от цГМФ, представляют собой важный путь для NO, чтобы контролировать свои молекулярные мишени. Эти пути в основном представлены S-нитрозилированием белков, ковалентным присоединением NO к тиоловой группе остатков цистеина (Cys) [78].Сообщалось о значительном снижении уровня S-нитрозилированных белков в гипоксическом сердце золотой рыбки по сравнению с нормоксическим аналогом. У млекопитающих значение дисрегуляции S-нитрозилирования белка коррелирует как с сердечными расстройствами [79], так и с защитными механизмами против развития дисфункции миокарда при стрессе [80]. Белки, подвергающиеся денитрозилированию в гипоксическом сердце золотых рыбок, и связанное с ними функциональное значение еще не идентифицированы.Однако разумно предположить, что это событие, активируя еще не определенные защитные программы, способствует сохранению функции миокарда в условиях гипоксии [75].
Интересно, что в гипоксическом сердце золотых рыбок молекулярные события генерации NO ниже по течению исключают участие цГМФ-зависимой передачи сигналов [75]. Пути, не зависящие от цГМФ, представляют собой важный путь для NO, чтобы контролировать свои молекулярные мишени. Эти пути в основном представлены S-нитрозилированием белков, ковалентным присоединением NO к тиоловой группе остатков цистеина (Cys) [78].Сообщалось о значительном снижении уровня S-нитрозилированных белков в гипоксическом сердце золотой рыбки по сравнению с нормоксическим аналогом. У млекопитающих значение дисрегуляции S-нитрозилирования белка коррелирует как с сердечными расстройствами [79], так и с защитными механизмами против развития дисфункции миокарда при стрессе [80]. Белки, подвергающиеся денитрозилированию в гипоксическом сердце золотых рыбок, и связанное с ними функциональное значение еще не идентифицированы.Однако разумно предположить, что это событие, активируя еще не определенные защитные программы, способствует сохранению функции миокарда в условиях гипоксии [75].
Известно, что в условиях гипоксии NO может определять нитрование белков. Он заключается в замещении, в основном под действием пероксинитрита (ОНОО —), нитрогруппы на остатки тирозина с образованием 3-нитротирозина [81]. Нитрование обычно связано с изменениями белкового катализа, белок-белкового взаимодействия и передачи сигналов тирозинкиназы [82].Однако зависимый от нитрования контроль окислительно-восстановительного гомеостаза присутствует и в нормально функционирующей сердечной мышце [83]. Интересно, что данные, полученные на гипоксическом сердце золотых рыбок, указывают на наличие индуцированного гипоксией нитрования, поскольку сообщалось об увеличении экспрессии Nox2, каталитической субъединицы НАДФН-оксидазы [75], и 3-нитротирозина [84]. Если дальнейшие данные подтвердят наличие нитрования, необходимо определить предполагаемые цели. На основании имеющейся информации можно предположить существование некоторых белков.Одним из них является насос SERCA2a, интегральный мембранный белок, контролирующий гомеостаз сердечного Ca 2+ путем активного транспорта иона в саркоплазматический ретикулум. Он восприимчив к нитрозативным и окислительным модификациям из-за присутствия нескольких остатков цистеина и тирозина [83,85]. Структурная близость к митохондриям подвергает насосы SERCA2a воздействию реактивных форм O 2 / азота, образующихся в качестве побочных продуктов окислительного фосфорилирования [86]. Нитротирозиновая модификация SERCA2a наблюдалась при нескольких патофизиологических состояниях [87], а нитрованный SERCA2a используется в качестве сердечного маркера нитративного стресса [83].Хотя прямых доказательств того, что SERCA2a вызывает нитрацию у рыб, нет, значительное снижение индуцированного гипоксией с течением времени увеличения производительности сердца золотых рыбок, наблюдаемое в условиях ингибирования SERCA2a [75], указывает на SERCA2a как на предполагаемую мишень нитрования в организме. гипоксическое сердце.
Он восприимчив к нитрозативным и окислительным модификациям из-за присутствия нескольких остатков цистеина и тирозина [83,85]. Структурная близость к митохондриям подвергает насосы SERCA2a воздействию реактивных форм O 2 / азота, образующихся в качестве побочных продуктов окислительного фосфорилирования [86]. Нитротирозиновая модификация SERCA2a наблюдалась при нескольких патофизиологических состояниях [87], а нитрованный SERCA2a используется в качестве сердечного маркера нитративного стресса [83].Хотя прямых доказательств того, что SERCA2a вызывает нитрацию у рыб, нет, значительное снижение индуцированного гипоксией с течением времени увеличения производительности сердца золотых рыбок, наблюдаемое в условиях ингибирования SERCA2a [75], указывает на SERCA2a как на предполагаемую мишень нитрования в организме. гипоксическое сердце.
В целом из имеющейся информации следует, что в сердце золотых рыбок NO активирует защитную программу, которая поддерживает работоспособность в условиях гипоксии. В соответствии с этим было обнаружено, что NO положительно модулирует кардиальные сарколеммальные КАТФ-каналы, ответ, который, как и КАТФ-зависимая защита, наблюдаемая в ишемизированном миокарде млекопитающих [88], может способствовать толерантности сердечной гипоксии у этого вида [8].Кроме того, гипоксическое сердце золотой рыбки также демонстрирует повышенную экспрессию индуцируемого гипоксией фактора α (HIF-1 α) [11]. У млекопитающих взаимодействие HIF1α/NO участвует в кардиозащитных реакциях, вызванных гипоксией. В условиях гипоксии HIF-1α активирует гены, важные для выживания клеток, включая NOS [89,90,91]; в то же время высокие концентрации NO (> 1 мкМ) стабилизируют HIF-1α, что приводит к увеличению димерной формы белка, который, связываясь с сайтами HRE, усиливает экспрессию гена NOS и, таким образом, образование NO [92].
В соответствии с этим было обнаружено, что NO положительно модулирует кардиальные сарколеммальные КАТФ-каналы, ответ, который, как и КАТФ-зависимая защита, наблюдаемая в ишемизированном миокарде млекопитающих [88], может способствовать толерантности сердечной гипоксии у этого вида [8].Кроме того, гипоксическое сердце золотой рыбки также демонстрирует повышенную экспрессию индуцируемого гипоксией фактора α (HIF-1 α) [11]. У млекопитающих взаимодействие HIF1α/NO участвует в кардиозащитных реакциях, вызванных гипоксией. В условиях гипоксии HIF-1α активирует гены, важные для выживания клеток, включая NOS [89,90,91]; в то же время высокие концентрации NO (> 1 мкМ) стабилизируют HIF-1α, что приводит к увеличению димерной формы белка, который, связываясь с сайтами HRE, усиливает экспрессию гена NOS и, таким образом, образование NO [92].
2.3. Температура
Водные эктотермы зависят от термической среды, которая регулирует скорость их метаболизма. Термическая толерантность ограничена способностью удовлетворять адекватные потребности O 2 тканей [93,94,95,96]. Помимо видов, обитающих в крайне стабильных условиях, многие рыбы регулярно сталкиваются с колебаниями температуры, связанными не только с онтогенетическими и/или сезонными изменениями, но и с суточными изменениями, особенно в мелководных водоемах. Тем не менее, их фенотипическая пластичность (развивающая или обратимая акклиматизация) позволяет компенсировать изменение пределов толерантности для оптимизации продуктивности при изменении температурных режимов [97,98,99,100,101].В то время как эвритермные рыбы, естественно подверженные большим изменениям температуры, вырабатывают стратегии акклиматизации для сохранения своей приспособленности, стенотермные виды демонстрируют специфические эволюционные адаптации за счет снижения пластичности.
Помимо видов, обитающих в крайне стабильных условиях, многие рыбы регулярно сталкиваются с колебаниями температуры, связанными не только с онтогенетическими и/или сезонными изменениями, но и с суточными изменениями, особенно в мелководных водоемах. Тем не менее, их фенотипическая пластичность (развивающая или обратимая акклиматизация) позволяет компенсировать изменение пределов толерантности для оптимизации продуктивности при изменении температурных режимов [97,98,99,100,101].В то время как эвритермные рыбы, естественно подверженные большим изменениям температуры, вырабатывают стратегии акклиматизации для сохранения своей приспособленности, стенотермные виды демонстрируют специфические эволюционные адаптации за счет снижения пластичности.
У многих эвритермных рыб изменения температуры существенно влияют на сердечную функцию, которую необходимо модулировать для обеспечения адекватного СО. Кроме того, верхняя термостойкость частично определяется способностью сердца обеспечивать адекватный системный O 2 доставка [102]. Это происходит за счет изменения частоты сердечных сокращений (ЧСС) больше, чем ударного объема [103, 104]. При резком повышении температуры ЧСС увеличивается, а затем снижается при температурах, предшествующих критическому тепловому максимуму [103, 104], и это ставит под угрозу сердечную функцию [102, 105, 106]. С другой стороны, при снижении температуры возникает брадикардия [107], и это связано с увеличением продолжительности диастолы для поддержания СВ за счет увеличения времени наполнения и с небольшими изменениями продолжительности систолы [108].Кроме того, длительные колебания температуры вызывают ремоделирование сердечной функции, но эффекты могут быть совершенно иными, чем наблюдаемые при острых изменениях ([107] и ссылки в ней). На самом деле, в то время как резкое повышение температуры обычно увеличивает скорость потенциала кардиостимулятора, более длительное воздействие более высокой температуры снижает внутреннюю ЧСС [109, 110]. Сброс частоты кардиостимулятора может происходить в разных временных масштабах и может зависеть от разных популяций, видов и экспериментальных условий [110, 111, 112]. Кроме того, на возбудимость миокарда влияет температура. При критически высоких температурах это объясняется дисбалансом входящих I Na и внешних I K1 токов [113, 114]. Изменения электрических свойств миокарда вызывают функциональную атриовентрикулярную блокаду с последующей брадикардией, ограниченной желудочком, который становится неспособным следовать синусовому ритму [114].
Это происходит за счет изменения частоты сердечных сокращений (ЧСС) больше, чем ударного объема [103, 104]. При резком повышении температуры ЧСС увеличивается, а затем снижается при температурах, предшествующих критическому тепловому максимуму [103, 104], и это ставит под угрозу сердечную функцию [102, 105, 106]. С другой стороны, при снижении температуры возникает брадикардия [107], и это связано с увеличением продолжительности диастолы для поддержания СВ за счет увеличения времени наполнения и с небольшими изменениями продолжительности систолы [108].Кроме того, длительные колебания температуры вызывают ремоделирование сердечной функции, но эффекты могут быть совершенно иными, чем наблюдаемые при острых изменениях ([107] и ссылки в ней). На самом деле, в то время как резкое повышение температуры обычно увеличивает скорость потенциала кардиостимулятора, более длительное воздействие более высокой температуры снижает внутреннюю ЧСС [109, 110]. Сброс частоты кардиостимулятора может происходить в разных временных масштабах и может зависеть от разных популяций, видов и экспериментальных условий [110, 111, 112]. Кроме того, на возбудимость миокарда влияет температура. При критически высоких температурах это объясняется дисбалансом входящих I Na и внешних I K1 токов [113, 114]. Изменения электрических свойств миокарда вызывают функциональную атриовентрикулярную блокаду с последующей брадикардией, ограниченной желудочком, который становится неспособным следовать синусовому ритму [114].
Ремоделирование сердца сопровождает реакцию сердца рыб на температурную акклиматизацию [107,115,116,117].Однако большие внутри- и межвидовые различия не позволяют составить общую картину, описывающую сердечную реакцию либо на острое воздействие тепла/холода, либо на акклиматизацию к температурным воздействиям. Уменьшение массы желудочков наблюдается при теплой акклиматизации с последующим снижением ударного объема и без изменений СВ, которое поддерживается за счет улучшения сократительной способности и более быстрого сокращения и расслабления желудочков [115]. И наоборот, холодовая акклиматизация вызывает гипертрофию сердца.В этом случае большая мышечная масса желудочка компенсирует снижение сократительной способности, поддерживая ударный объем и, следовательно, СВ при более низких температурах [107, 115, 116].
И наоборот, холодовая акклиматизация вызывает гипертрофию сердца.В этом случае большая мышечная масса желудочка компенсирует снижение сократительной способности, поддерживая ударный объем и, следовательно, СВ при более низких температурах [107, 115, 116].
В отличие от эвритермных видов, стенотермные рыбы практически не переносят тепловые нагрузки. Это случай антарктических костистых рыб Channichthyidae, которые живут в чрезвычайно стабильных, холодных и насыщенных кислородом антарктических водах [118, 119]. Некоторые из них уникальны среди взрослых позвоночных, так как у них отсутствует гемоглобин (Hb; [120]), а у некоторых видов также Mb [121].Это компенсируется экстенсивным ремоделированием сердечно-сосудистой системы, таким как гиперволемия, низкая вязкость крови, крупные капилляры, кардиомегалия и высокий кровоток с низким системным давлением и системным сопротивлением ([119] и ссылки в ней). Возможно, что отсутствие дыхательных пигментов может способствовать умеренной термической пластичности, что делает этих костистых рыб уязвимыми к кратковременным экстремальным колебаниям температуры, а также к долговременному потеплению климата [122,123]. Однако недавние сравнительные исследования, проведенные на нескольких антарктических костистых с дыхательными пигментами и без них, предполагают некоторую степень аккомодации сердца к колебаниям температуры, особенно у видов, экспрессирующих как Hb, так и Mb [124].
Однако недавние сравнительные исследования, проведенные на нескольких антарктических костистых с дыхательными пигментами и без них, предполагают некоторую степень аккомодации сердца к колебаниям температуры, особенно у видов, экспрессирующих как Hb, так и Mb [124].
2.4. Система NOS/NO в сердце рыб при температурных воздействиях
У рыб система NOS/NO играет роль в регуляции сердечной функции у видов, адаптированных как к умеренному, так и к экстремальному тепловому режиму, а также у животных с разной толерантностью к тепловому стрессу.
У эвритермального угря A. anguilla NOS/NO-зависимая модуляция реакции Франка-Старлинга [32,34] нарушается при изменении температуры [125]. Амелио с соавторами наблюдали, что положительная модуляция реакции Франка-Старлинга, вызванная внутрисердечным высвобождением NO, исчезает при резком воздействии на животных температур ниже или выше акклиматизационной, как в случае весеннего, так и зимнего типа (температура акклиматизации). : 20 °C и 10 °C соответственно).Эти эффекты сопровождаются снижением уровней экспрессии NOS и pAkt, что свидетельствует о том, что продукция NO через ось Akt/NOS зависит от температуры [125]. Авторы предполагают, что отмена нитрергической модуляции реакции Франка-Старлинга при термических нагрузках может включать протеинкиназы, предполагая, что Akt является чувствительным к температуре элементом в NO-генерирующем трансдукционном каскаде [125].
: 20 °C и 10 °C соответственно).Эти эффекты сопровождаются снижением уровней экспрессии NOS и pAkt, что свидетельствует о том, что продукция NO через ось Akt/NOS зависит от температуры [125]. Авторы предполагают, что отмена нитрергической модуляции реакции Франка-Старлинга при термических нагрузках может включать протеинкиназы, предполагая, что Akt является чувствительным к температуре элементом в NO-генерирующем трансдукционном каскаде [125].
Другим примером являются лососевые. У эвритермального атлантического лосося длительное воздействие повышения температуры сопровождается повышенной экспрессией iNOS как в компактном, так и в губчатом желудочковом миокарде, что свидетельствует об усиленной продукции NO.В то же время увеличивается и экспрессия VEGF [126]. Это интересно, так как два эффекта, если рассматривать их вместе, могут требовать потенцирования кровоснабжения миокарда за счет увеличения васкуляризации (через VEGF) и/или за счет расширения сосудов (через NO). На самом деле известно, что у лососевых NO вызывает вазодилатацию и снижает коронарное сопротивление [19], тем самым способствуя компенсации повышенного потребления O 2 при повышенной температуре.
Взаимосвязь между колебаниями температуры и нитрергическим контролем сердца может иметь большое значение для рыб, живущих при экстремальных температурах.К сожалению, этот аспект остается неизученным, хотя имеющаяся на сегодняшний день информация указывает на то, что система NOS/NO играет роль в модуляции основных функций сердца у этих животных.
У антарктических костистых рыб функциональные NOS присутствуют в сердце C. aceratus без гемоглобина и C. hamatus и краснокровных T. bernacchii. eNOS-подобный фермент в основном присутствует в лакунах губчатого желудочка, в то время как iNOS базально экспрессируется в цитоплазме миокардиоцитов [27,28].Несмотря на сходное распределение, физико-фармакологические исследования показывают, что NO по-разному влияет на сократительную способность трех видов [28]. Фактически эндогенный NO (введение L-аргинина) снижает сократительную способность T. bernacchii , в отличие от стимулирующего эффекта, наблюдаемого у двух видов ледяных рыб. Кроме того, в то время как у C. hamatus NO-индуцированные эффекты зависят от цГМФ, у T. bernacchii и C. aceratus эти эффекты не зависят от цГМФ.Авторы предполагают, что в отсутствие дыхательных пигментов потеря активности NO-оксигеназы, связанная с Hb/Mb, и последующее повышение уровня NO могут объяснить наблюдаемые различия [28]. Кроме того, более высокое количество NO у ледяной рыбы может иметь значение для уравновешивания адренергической стимуляции сократительной способности in vivo, которая является частью теплового ответа сердца этих костистых рыб [124]. Приветствуются дальнейшие исследования, чтобы понять, может ли отсутствие дыхательных пигментов, связанное с повышенной доступностью NO в сердце, быть полезным или вредным для устойчивости этих рыб к длительным температурным испытаниям.
Кроме того, в то время как у C. hamatus NO-индуцированные эффекты зависят от цГМФ, у T. bernacchii и C. aceratus эти эффекты не зависят от цГМФ.Авторы предполагают, что в отсутствие дыхательных пигментов потеря активности NO-оксигеназы, связанная с Hb/Mb, и последующее повышение уровня NO могут объяснить наблюдаемые различия [28]. Кроме того, более высокое количество NO у ледяной рыбы может иметь значение для уравновешивания адренергической стимуляции сократительной способности in vivo, которая является частью теплового ответа сердца этих костистых рыб [124]. Приветствуются дальнейшие исследования, чтобы понять, может ли отсутствие дыхательных пигментов, связанное с повышенной доступностью NO в сердце, быть полезным или вредным для устойчивости этих рыб к длительным температурным испытаниям.
Тропические двоякодышащие рыбы представляют собой своеобразный модельный организм, поскольку в теплое время года у них происходит аэстивация, метаболическая адаптация, связанная с функциональными изменениями в тканях и органах, включая сердце, почки, жабры, легкие и скелетные мышцы [10,29,127,128,129]. Очень интересным аспектом двоякодышащих рыб является способность миокарда обеспечивать сократимость во время теплового отдыха, поддерживая соответствующее кровоснабжение всего организма. Сердце двоякодышащих рыб, наблюдаемое у Protopterus dolloi [29] и Protopterus annectens [10], экспрессирует ферменты NOS, и эта экспрессия увеличивается при эстивации [29].Было высказано предположение, что последующее усиленное высвобождение NO защищает сердце, поддерживая сердечную биоэнергетику при метаболической депрессии и сниженном потреблении миокардом O 2 [10, 29, 128].
Очень интересным аспектом двоякодышащих рыб является способность миокарда обеспечивать сократимость во время теплового отдыха, поддерживая соответствующее кровоснабжение всего организма. Сердце двоякодышащих рыб, наблюдаемое у Protopterus dolloi [29] и Protopterus annectens [10], экспрессирует ферменты NOS, и эта экспрессия увеличивается при эстивации [29].Было высказано предположение, что последующее усиленное высвобождение NO защищает сердце, поддерживая сердечную биоэнергетику при метаболической депрессии и сниженном потреблении миокардом O 2 [10, 29, 128].
2.5. От одного стресса к множественному стрессу: в сердце рыбы есть место для азотергической системы
Сердечная функция постоянно подвергается сомнению из-за конвергенции множественных стрессов. У животных, обитающих в водной среде, О 2 чрезвычайно часты ограничения и перепады температуры.Однако перекрестное взаимодействие между этими двумя переменными в сердце почти не исследовано.
У рыб доступность O 2 считается главным определяющим фактором термоустойчивости, за исключениями, которые, возможно, связаны со специфическими адаптациями и/или ролью дополнительных внутренних и/или внешних ограничений [130]. Совсем недавно были проведены эксперименты на оборудованном in situ сердце устойчивой к гипоксии угольной рыбы ( Anoplopoma fimbria ( A.fimbria )) попытались проанализировать перекрестную толерантность между двумя связанными событиями ограничения O 2 и повышенной температуры. Результаты показывают, что предварительное воздействие низкого O 2 с последующим согреванием делает сердце неспособным увеличивать ЧСС и СВ. Авторы предполагают роль нервной системы и выдвигают гипотезу, что гипоксия может серьезно ограничивать сердечный ответ на тепловые нагрузки [1]. 96].
Кроме того, мало внимания уделялось роли системы NOS/NO, когда сердце рыбы подвергается одновременному воздействию температуры и O 2 вариаций, так что на данный момент можно сделать любой вывод. только выводное.Функциональные данные по желудочковым полоскам форели ( O. mykiss) показали, что благотворное влияние NO на работу сердца в условиях гипоксии снижается или даже обращается вспять, если ЧСС увеличивается [31]. Интересно, что, как показано выше, ускорение является типичной реакцией сердца рыб на повышение температуры, что имеет решающее значение для соответствия насосной функции потребностям организма [103, 131]. Соответственно, можно предположить, что если рыба подвергается как повышению температуры, так и ограничению O 2 , усиление ЧСС может усугубить последствия потери NO-зависимой защиты на сократительную способность.
только выводное.Функциональные данные по желудочковым полоскам форели ( O. mykiss) показали, что благотворное влияние NO на работу сердца в условиях гипоксии снижается или даже обращается вспять, если ЧСС увеличивается [31]. Интересно, что, как показано выше, ускорение является типичной реакцией сердца рыб на повышение температуры, что имеет решающее значение для соответствия насосной функции потребностям организма [103, 131]. Соответственно, можно предположить, что если рыба подвергается как повышению температуры, так и ограничению O 2 , усиление ЧСС может усугубить последствия потери NO-зависимой защиты на сократительную способность.
На субклеточном уровне как температурные изменения, так и гипоксия сходятся на митохондриях, которые чувствительны к температуре и O 2 -чувствительным. Это важно для рыб, которые, будучи экзотермами, нуждаются в оптимизации аэробного метаболизма при изменении температуры [132, 133]. При резком повышении температуры до температурного предела функция митохондрий сердца рыб изменяется с точки зрения скорости утечки протонов, окислительного фосфорилирования, целостности мембран, функции белковых комплексов и окислительного дисбаланса из-за увеличения продукции АФК [132, 134, 135, 136]. Однако, как сообщалось у атлантического лосося ( S. salar ), акклиматизация к потеплению (20 °C, близко к верхней температурной толерантности вида) смягчает нарушение сердечной митохондриальной функции, предполагая пластичность, которая может быть полезной для теплового воздействия. толерантность. Кроме того, акклиматизация к теплу снижает NO-зависимое ингибирование функции сердечных митохондрий [137, 138]. Это может лучше соответствовать повышенным метаболическим потребностям, связанным с тепловым стрессом, хотя неизвестно, полезно ли это во время акклиматизации к температурам, близким к верхней температурной толерантности [138].
Однако, как сообщалось у атлантического лосося ( S. salar ), акклиматизация к потеплению (20 °C, близко к верхней температурной толерантности вида) смягчает нарушение сердечной митохондриальной функции, предполагая пластичность, которая может быть полезной для теплового воздействия. толерантность. Кроме того, акклиматизация к теплу снижает NO-зависимое ингибирование функции сердечных митохондрий [137, 138]. Это может лучше соответствовать повышенным метаболическим потребностям, связанным с тепловым стрессом, хотя неизвестно, полезно ли это во время акклиматизации к температурам, близким к верхней температурной толерантности [138].
В нескольких исследованиях на рыбах изучалось влияние ограничения O 2 на митохондриальные функции и последствия для работы сердца. Были получены противоречивые данные, возможно, в зависимости от экспериментальных условий и/или видовой устойчивости к гипоксии [139,140]. Точно так же роль нитрергической системы почти игнорировалась, за исключением данных, полученных у угольной рыбы A. fimbria , у которых изучалось влияние NO на митохондриальное дыхание вместе с митохондриальной чувствительностью к NO в отношении O 2 доступность [141].Установлено, что длительное воздействие гипоксии на животных повышает чувствительность митохондрий сердца к ингибированию NO. Однако, как также отмечают авторы, неясно, можно ли считать это общей реакцией или своеобразной чертой этого вида, характеризующейся высокой толерантностью к гипоксии [141]. Насколько нам известно, нет информации об участии нитрергической системы в модуляции сердечной митохондриальной функции рыб, когда тепловые приращения и гипоксия конвергируют у животных.
fimbria , у которых изучалось влияние NO на митохондриальное дыхание вместе с митохондриальной чувствительностью к NO в отношении O 2 доступность [141].Установлено, что длительное воздействие гипоксии на животных повышает чувствительность митохондрий сердца к ингибированию NO. Однако, как также отмечают авторы, неясно, можно ли считать это общей реакцией или своеобразной чертой этого вида, характеризующейся высокой толерантностью к гипоксии [141]. Насколько нам известно, нет информации об участии нитрергической системы в модуляции сердечной митохондриальной функции рыб, когда тепловые приращения и гипоксия конвергируют у животных.
Пути NOS/NO, активируемые в сердце рыб в ответ на гипоксию и температурный стресс, обобщены в .
Схематический обзор NOS/NO-опосредованных внутриклеточных путей, активируемых в кардиомиоцитах рыб (MYO) при базальном (черные стрелки), гипоксическом (фиолетовые стрелки) и термическом стрессе (синяя и красная стрелки). Подробности смотрите в тексте.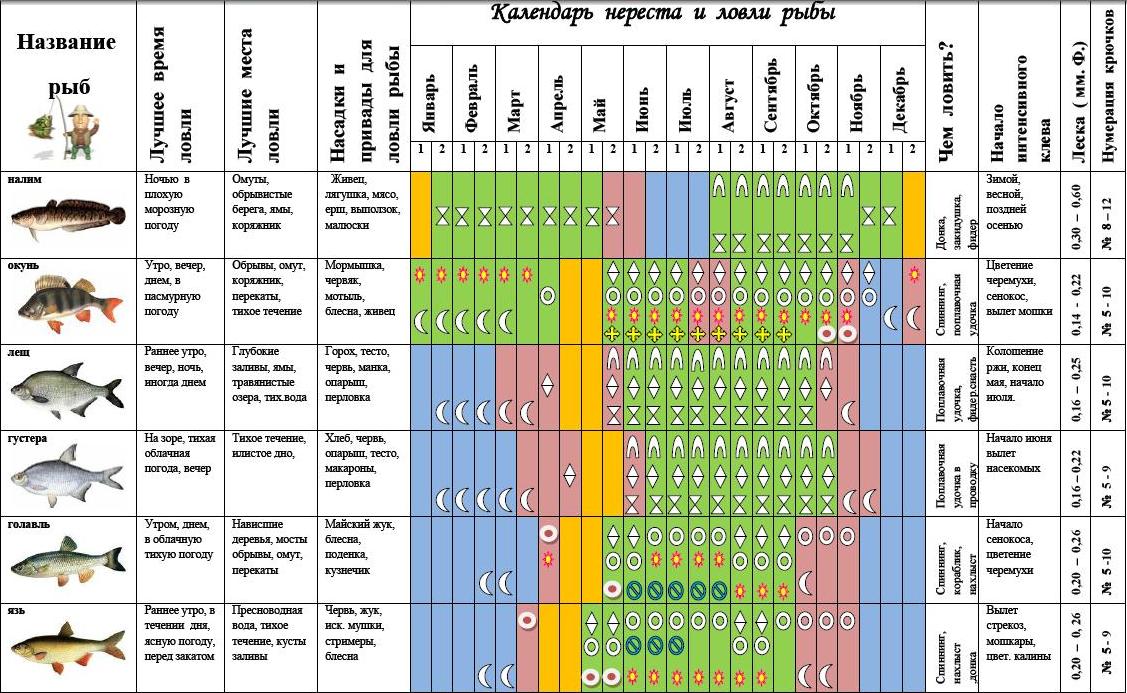 AC, аденлилатциклаза; AKT, серин/треонинкиназа 1; β3-AR, бета-3-адренорецептор; цАМФ, циклический аденозинмонофосфат; цГМФ, циклический гуанозинмонофосфат; EE, эндокардиальный эндотелий; HIF-1α, индуцируемый гипоксией фактор-1α; Mb, миоглобин; NO, оксид азота; NOS, синтаза оксида азота; № 2 – , нитрит; NOX2, НАДФН-оксидаза; NR, нитритредуктаза; ONOO – , пероксинитрит; PI-3K, фосфатидилинозитол-3-киназа; PKA, протеинкиназа А; PKG, протеинкиназа G; PLB, фосфоламбан; RyR, рианодиновый рецептор; SERCA, саркоэндоплазматический ретикулум Са 2+ -АТФазный насос; рГЦ, растворимая гуанилатциклаза; SNO, S-нитрозилирование; SR, саркоплазматический ретикулум; VEGF, фактор роста эндотелия сосудов.
AC, аденлилатциклаза; AKT, серин/треонинкиназа 1; β3-AR, бета-3-адренорецептор; цАМФ, циклический аденозинмонофосфат; цГМФ, циклический гуанозинмонофосфат; EE, эндокардиальный эндотелий; HIF-1α, индуцируемый гипоксией фактор-1α; Mb, миоглобин; NO, оксид азота; NOS, синтаза оксида азота; № 2 – , нитрит; NOX2, НАДФН-оксидаза; NR, нитритредуктаза; ONOO – , пероксинитрит; PI-3K, фосфатидилинозитол-3-киназа; PKA, протеинкиназа А; PKG, протеинкиназа G; PLB, фосфоламбан; RyR, рианодиновый рецептор; SERCA, саркоэндоплазматический ретикулум Са 2+ -АТФазный насос; рГЦ, растворимая гуанилатциклаза; SNO, S-нитрозилирование; SR, саркоплазматический ретикулум; VEGF, фактор роста эндотелия сосудов.
3. Выше и ниже по течению системы NOS/NO: AMP-активированная протеинкиназа (AMPK) как кандидат в рыбе
Несмотря на растущие данные, подтверждающие, что NO является ключевым медиатором сердечной реакции рыб на O 2 и температурный стресс, необходимы дальнейшие исследования, чтобы обнаружить полные молекулярные сети, которые организованы этим газотрансмиттером. Особого внимания заслуживает роль, которую играют киназы. Эти ферменты, в значительной степени экспрессируемые в клетках сердца и стратегически расположенные во внутриклеточных сигнальных путях, контролируют функцию белка и, следовательно, большое количество клеточных активностей.У млекопитающих хорошо оценена важность кардиального «кинома» (подробнее см. [142]). Некоторые из этих ферментов имеют решающее значение для ответа миокарда на стресс. Примерами являются киназы, относящиеся к каскадам RISK и SAFE, роль которых является фундаментальной для защиты миокарда от проблем, вызванных ишемией/реперфузией [143]. У рыб кардиальный кином в настоящее время недостаточно изучен. Однако на основании имеющихся данных, в том числе обобщенных выше, и данных по млекопитающим легко предположить вклад критических киназ в формирование адаптивного ответа на стресс.Вместе с PKA, PKG, PI3K, Akt и другими AMPK представляет собой интересного кандидата. Это чувствительная серин-треонинкиназа, состоящая из трех субъединиц (каталитическая α-субъединица 63 кДа, регуляторная β-субъединица 43 кДа и γ-субъединица 38 кДа), кодируемых разными генами, по-разному экспрессирующимися в клетках и тканях. Его основная роль заключается в поддержании клеточного гомеостаза по отношению к энергетическому балансу [144,145]. Высокие уровни AMP, возникающие, когда потребление АТФ превышает производство, активируют киназу, что приводит к общему переходу от анаболизма к катаболизму, модуляции экспрессии генов и белков, посттрансляционной модификации метаболических ферментов и биогенезу митохондрий [146,147]. .Активность AMPK также связана с метаболическим окислительно-восстановительным и окислительным равновесием, хотя окончательные модели причинно-следственных связей все еще находятся в стадии разработки. Было высказано предположение, что киназа либо инактивируется окислительным стрессом [148], либо стимулируется воздействием H 2 O 2 и NO [149]. В наши цели не входит подробное описание этой богатой молекулярной сети, и читателю предлагается обратиться к множеству превосходных и исчерпывающих обзоров для получения дополнительной информации.
Его основная роль заключается в поддержании клеточного гомеостаза по отношению к энергетическому балансу [144,145]. Высокие уровни AMP, возникающие, когда потребление АТФ превышает производство, активируют киназу, что приводит к общему переходу от анаболизма к катаболизму, модуляции экспрессии генов и белков, посттрансляционной модификации метаболических ферментов и биогенезу митохондрий [146,147]. .Активность AMPK также связана с метаболическим окислительно-восстановительным и окислительным равновесием, хотя окончательные модели причинно-следственных связей все еще находятся в стадии разработки. Было высказано предположение, что киназа либо инактивируется окислительным стрессом [148], либо стимулируется воздействием H 2 O 2 и NO [149]. В наши цели не входит подробное описание этой богатой молекулярной сети, и читателю предлагается обратиться к множеству превосходных и исчерпывающих обзоров для получения дополнительной информации.
В контексте этого обзора аспект, который необходимо выделить, — это взаимосвязь между AMPK и нитрергической системой. Киназа расположена выше ферментов NOS. В культивируемых кардиомиоцитах млекопитающих AMPK активирует eNOS и nNOS путем фосфорилирования, тем самым способствуя продукции NO [150]. В то же время AMPK-зависимый путь активирует Akt, тем самым косвенно модулируя то, что в кардиомиоцитах участвует в фосфорилировании eNOS [151].
Киназа расположена выше ферментов NOS. В культивируемых кардиомиоцитах млекопитающих AMPK активирует eNOS и nNOS путем фосфорилирования, тем самым способствуя продукции NO [150]. В то же время AMPK-зависимый путь активирует Akt, тем самым косвенно модулируя то, что в кардиомиоцитах участвует в фосфорилировании eNOS [151].
В сердце взаимосвязь между AMPK и нитрергической системой имеет большое значение, особенно при стрессе. AMPK активируется при ишемии-реперфузии, повышенной рабочей нагрузке и нарушении усвоения глюкозы, при всех состояниях, требующих снижения O 2 и АТФ и нарушения окислительно-нитрозативного равновесия [152, 153, 154].Примечательно, что сама AMPK регулируется NO, который действует как эндогенный активатор киназы. В нормальных условиях NO стимулирует AMPK через путь sGC/cGMP/Ca 2+ /CaMKβ, тогда как при стрессе киназа активируется сверхпродуцируемым пероксинитритом через каскад PKC/LKB1 [149]. Значение этой положительной обратной связи между AMPK и нитрергической системой, а также механизмы реципрокного контроля все еще являются предметом активных дискуссий у млекопитающих.
Фундаментальное значение AMPK в клеточном гомеостазе однозначно подтверждается ее присутствием на протяжении всей эволюции живых организмов, от амебозойных до млекопитающих и даже у растений [155,156].У костистых рыб, в соответствии со случаем дупликации всего генома [157], субъединицы AMPK, кодирующие гены, расширяются, так что, например, у рыбок данио присутствует 10 субъединиц AMPK [155,156]. Это усиливает функции фермента и усложняет молекулярные исследования, проводимые с помощью экспериментальных схем и инструментов (например, антител), созданных для исследований на млекопитающих.
До сих пор биологическая роль фермента изучалась у нескольких видов костистых костей в связи с различными проблемами.Молекулярные и функциональные данные, полученные у устойчивых к гипоксии/аноксии карасей и золотых рыбок [158,159], позволяют предположить, что киназа чувствительна к восстановлению O 2 тканеспецифическим образом. У золотых рыбок, подвергшихся 12-часовой тяжелой гипоксии, AMPK активируется в печени и, по-видимому, не активируется в других тканях, включая сердце, мозг и жабры [159]. На данный момент неясно, является ли отсутствие активации ферментов одним из физиологических компенсаторных механизмов, предотвращающих снижение энергии при низком O 2 , таких как регионарные сбросы крови и/или активация альтернативных метаболических путей, способных поддерживать биоэнергетику клетки. 60].В то же время на имеющиеся данные может влиять экспериментальный контекст. Например, у золотых рыбок короткого времени воздействия и частичной доступности O 2 может быть недостаточно, чтобы вызвать активацию фермента [159]. В соответствии с этим у карася длительное воздействие (до 7 дней) тяжелой аноксии, но не гипоксии, сопровождается повышенной активацией (т. е. фосфорилированием) AMPK в сердце. Это предполагает, что киназа может находиться в состоянии покоя до тех пор, пока не будет достигнута полная аноксия [160].Кроме того, это событие является обратимым, поскольку после реоксигенации уровень фосфорилированного фермента возвращается к преаноксическому уровню, что указывает на адаптивную роль [158].
На данный момент неясно, является ли отсутствие активации ферментов одним из физиологических компенсаторных механизмов, предотвращающих снижение энергии при низком O 2 , таких как регионарные сбросы крови и/или активация альтернативных метаболических путей, способных поддерживать биоэнергетику клетки. 60].В то же время на имеющиеся данные может влиять экспериментальный контекст. Например, у золотых рыбок короткого времени воздействия и частичной доступности O 2 может быть недостаточно, чтобы вызвать активацию фермента [159]. В соответствии с этим у карася длительное воздействие (до 7 дней) тяжелой аноксии, но не гипоксии, сопровождается повышенной активацией (т. е. фосфорилированием) AMPK в сердце. Это предполагает, что киназа может находиться в состоянии покоя до тех пор, пока не будет достигнута полная аноксия [160].Кроме того, это событие является обратимым, поскольку после реоксигенации уровень фосфорилированного фермента возвращается к преаноксическому уровню, что указывает на адаптивную роль [158]. Это может быть выгодно для устойчивых к гипоксии видов, поскольку позволяет повысить толерантность к гипоксии до того, как будут задействованы AMPK-опосредованные метаболические корректировки. Таким образом, синтез белка и другие анаболические пути, подавляемые AMPK, продолжают функционировать в условиях гипоксии наряду со способностью преимущественно направлять кровоток к органам, подверженным риску (например,г., мозг и сердце) [160].
Это может быть выгодно для устойчивых к гипоксии видов, поскольку позволяет повысить толерантность к гипоксии до того, как будут задействованы AMPK-опосредованные метаболические корректировки. Таким образом, синтез белка и другие анаболические пути, подавляемые AMPK, продолжают функционировать в условиях гипоксии наряду со способностью преимущественно направлять кровоток к органам, подверженным риску (например,г., мозг и сердце) [160].
В соответствии с ролью мультистрессор-зависимой киназы у рыб функция AMPK связана с температурными проблемами, хотя роль киназы до сих пор неясна. У лососевых ( O. mykiss и O. kisutch ) фосфорилирование AMPK коррелирует с оптимальной для аэробных условий температурой, измеряемой по максимальному ЧСС [161]. Недавние наблюдения за сердцем оливковой камбалы, Paralichthys olivaceus , показывают, что холодовой стресс активирует AMPK вместе с его вышестоящими модуляторами (LKB1 и CAMKK) и нижестоящими мишенями (SITR1, FOXO1A и TFAM), и это положительно влияет на рыбу. адаптивная реакция на холод [162].Однако Нильссон и его сотрудники [163] не сообщили об изменениях в фосфорилировании AMPK в сердце карася, акклиматизированного к температурам (4, 10 и 20 °C), естественным для этого вида, предполагая, что киназа остается в состоянии покоя в течение адаптивный температурный диапазон животного.
адаптивная реакция на холод [162].Однако Нильссон и его сотрудники [163] не сообщили об изменениях в фосфорилировании AMPK в сердце карася, акклиматизированного к температурам (4, 10 и 20 °C), естественным для этого вида, предполагая, что киназа остается в состоянии покоя в течение адаптивный температурный диапазон животного.
Насколько нам известно, взаимодействию между AMPK и нитрергической системой в сердце рыб в отношении стресса, одиночного или множественного, уделялось мало или даже не уделялось внимания.Однако проиллюстрированные выше данные ясно показывают, что оба присутствуют и активны в сердце рыбы и участвуют в адаптивной реакции на O 2 и изменения температуры. Возможность того, что чрезвычайная гибкость сердца некоторых видов рыб, как в случае карповых, может использовать сигнальные пути, которые конвергируют как в системе NOS/NO, так и в AMPK, интригует и заслуживает внимания.
Карибская торговля и сети (Служба национальных парков США)
Изделия с зонированной резьбой (ZIC), ранний саладоид. кошка. № 2465; б: кот. № 2466; в: кот. № 2478; д: кот. № 2471; е: кот. № 2462. Все осколки либо из стоянок Сент-Джорджес, либо из Ричмонда. Коллекция Фолмера Андерсона, Национальный исторический памятник Кристианстед
кошка. № 2465; б: кот. № 2466; в: кот. № 2478; д: кот. № 2471; е: кот. № 2462. Все осколки либо из стоянок Сент-Джорджес, либо из Ричмонда. Коллекция Фолмера Андерсона, Национальный исторический памятник Кристианстед(фото NPS)
Все протестированные образцы сковородок принадлежали к Группе 2, как и изделия ZIC, и все, кроме одного, кусочки красного на ободке или красного расписные изделия периода Корал-Бей-Лонгфорд (около 400-600 гг. Н.э.).
Затем результаты сравнивались с базой данных карибской керамики (также хранящейся в MURR).Было обнаружено, что несколько черепков из Доминиканской Республики и Пуэрто-Рико, вероятно, были сделаны на острове Санта-Крус. Другими словами, они были членами группы 2.
Группа 1 была представлена тремя черепками из Солт-Ривер и двумя черепками из стоянки Сент-Джорджес, одна из которых представляла собой светлую известняковую посуду, а другая — очищенную WOR-изделию. Эти результаты также были сопоставлены с панкарибской базой данных доисторической керамики MURR. Из-за сильного совпадения между этими образцами Группы 1 и сильной группировкой черепков, перечисленных в базе данных как сделанных в Пуэрто-Рико, вполне вероятно, что эта керамика Группы 1 из Св.Croix также были сделаны в Пуэрто-Рико.
Остальные черепки и глины не могли быть отнесены ни к одной группе, и не было никаких связей между этими образцами и образцами с других островов, перечисленных в базе данных MURR. Эти образцы включали окрашенный в коричневый и белый цвет черепок с участка Prosperity, один образец рафинированной посуды WOR (участок Prosperity) и черепок с оранжевой пленкой и красной окраской, очерченной надрезами (участок Виндзор). Вполне возможно, что эти черепки могли быть сделаны из глины, полученной из неизвестного источника за пределами острова, или они могли быть сделаны из Св.Глины Croix, образцы которых не были отобраны.
Часть II. Каменный кабинет
Вторая часть посвящена перевозке полированных каменных кельтов или топоров и небольших каменных украшений, вырезанных в форме животных, в основном лягушек и других рептилий или амфибийных существ (рис. 5–7). Эти каменные артефакты являются наиболее цитируемым свидетельством существования древних сетей межостровного обмена на большие расстояния в Карибском море. Несколько стилей этих украшений были обнаружены в местах по всему региону, которые соответствуют исследованиям, проведенным с подобными украшениями и сырьем (Boomert 1987; Cody 1990, 1991; Crock and Bartone 1998; Kelker 1985; Knippenberg 2006; McGinnis 1997; Narganes Storde 1995a). , 1995b; Родригес, 1991; Уоттерс и Скаглион, 1984).Неизвестно, был ли перемещен необработанный, нетронутый камень или готовые изделия. Тщательно отполированные топоры и кельты из «зеленого камня» без признаков износа в результате использования в качестве инструментов были обнаружены в нескольких местах по всей Санта-Крус, включая Аклис, Джудит Фэнси, Просперити, Ричмонд, Сент-Джорджес и Солт-Ривер. Они также были обнаружены во многих других местах Карибского моря. Их полированная, безупречная поверхность является возможным свидетельством их использования в качестве ценных, важных символов силы и знания.
5–7). Эти каменные артефакты являются наиболее цитируемым свидетельством существования древних сетей межостровного обмена на большие расстояния в Карибском море. Несколько стилей этих украшений были обнаружены в местах по всему региону, которые соответствуют исследованиям, проведенным с подобными украшениями и сырьем (Boomert 1987; Cody 1990, 1991; Crock and Bartone 1998; Kelker 1985; Knippenberg 2006; McGinnis 1997; Narganes Storde 1995a). , 1995b; Родригес, 1991; Уоттерс и Скаглион, 1984).Неизвестно, был ли перемещен необработанный, нетронутый камень или готовые изделия. Тщательно отполированные топоры и кельты из «зеленого камня» без признаков износа в результате использования в качестве инструментов были обнаружены в нескольких местах по всей Санта-Крус, включая Аклис, Джудит Фэнси, Просперити, Ричмонд, Сент-Джорджес и Солт-Ривер. Они также были обнаружены во многих других местах Карибского моря. Их полированная, безупречная поверхность является возможным свидетельством их использования в качестве ценных, важных символов силы и знания. Многие из этих полированных топоров и кельтов были сделаны из различных «зеленокаменных» материалов, которые напоминают светло-зеленые жадеиты (Harlow et al. 2006).
Многие из этих полированных топоров и кельтов были сделаны из различных «зеленокаменных» материалов, которые напоминают светло-зеленые жадеиты (Harlow et al. 2006).
Лягушка и другие украшения в стиле амфибий (или «батрахиан») были подвешены в одном из двух стилей: в стиле муиракита (амазонский) с отверстиями для подвески, просверленными по бокам и сзади, чтобы их нельзя было увидеть спереди, и более простой стиль, в котором отверстия просверливаются спереди назад. Также были протестированы подвески других более простых стилей, сделанные из материалов, напоминающих бирюзу, малахит и амазонит.В общей сложности 39 артефактов из коллекции Фолмера Андерсена были отправлены в Департамент наук о Земле и планетах Американского музея естественной истории (Нью-Йорк) для идентификации с помощью сканирующего электронного микроскопа (СЭМ) с детектором обратно рассеянных электронов ( BSE) и энергодисперсионный рентгеноспектрометрический анализатор (ЭДС) PGT-Imix.
Анализ с использованием электронного микрозонда определил концентрации первичных элементов, составляющих минералы. Протестированные артефакты включали девять каменных кельтов и 16 резных каменных украшений, в том числе шесть лягушек или «батрахианских» стилей и десять других типов (стиль гусеницы и простые бусы) (рис. 8).Также на испытания были представлены две «зеленокаменные» отщепы и 12 кусков необработанной «зеленокаменной» и «бирюзовой». Было обнаружено, что многие каменные топоры и кельты, скорее всего, были сделаны из жадеита, доступного только в одном месте, в долине реки Мотагуа, Гватемала. Это место было источником многих нефритов, используемых майя; Долина Мотагуа была заселена с позднего доклассического периода (ок. 400 г. до н.э. — 200 г. н.э.) до позднего постклассического периода (ок. 900-1500 гг. Н.э.), при этом несколько участков служили производственными центрами или мастерскими.
Протестированные артефакты включали девять каменных кельтов и 16 резных каменных украшений, в том числе шесть лягушек или «батрахианских» стилей и десять других типов (стиль гусеницы и простые бусы) (рис. 8).Также на испытания были представлены две «зеленокаменные» отщепы и 12 кусков необработанной «зеленокаменной» и «бирюзовой». Было обнаружено, что многие каменные топоры и кельты, скорее всего, были сделаны из жадеита, доступного только в одном месте, в долине реки Мотагуа, Гватемала. Это место было источником многих нефритов, используемых майя; Долина Мотагуа была заселена с позднего доклассического периода (ок. 400 г. до н.э. — 200 г. н.э.) до позднего постклассического периода (ок. 900-1500 гг. Н.э.), при этом несколько участков служили производственными центрами или мастерскими.
Результаты этого анализа аналогичны результатам недавних исследований жадеитовых топоров, извлеченных из раскопок на острове Антигуа, где источник также был идентифицирован как Мотагуа (Harlow et al. 2006; Murphy et al. 2000). Эти исследования доказывают, что древние саладоиды, которые мигрировали и поселились на островах Карибского моря почти 2000 лет назад, имели какие-то контакты с обществами в регионах за пределами тех, которые традиционно ассоциировались с саладоидным культурным регионом (т.е. Карибский бассейн и нижняя часть долины реки Ориноко). С другой стороны, украшения из нефрита были вырезаны из различных материалов, обычно называемых социальными нефритами, а именно из нефрита, серпентина, хризопраза, альбитита и кварцитов, таких как фуксит. Многие из этих социальных нефритов могли быть добыты на соседних островах. Однако на данный момент источники этих артефактов неизвестны. «Бирюзовые» предметы были идентифицированы как малахит и псевдомалахит с примесью бирюзы. Малахит и бирюза образуются в сухом климате, выветриваются из медных руд и часто связаны с отложениями известняка (Джордж Харлоу, личное сообщение, 2008 г.).Месторождения меди, связанные с дуговым вулканизмом, расположены на Пуэрто-Рико, Виргинских островах (Сент-Джон) и Малых Антильских островах, и Кокс и Бриггс (1973: 3) отметили, что все месторождения меди на Пуэрто-Рико «содержат некоторое количество малахита или азурита.
2006; Murphy et al. 2000). Эти исследования доказывают, что древние саладоиды, которые мигрировали и поселились на островах Карибского моря почти 2000 лет назад, имели какие-то контакты с обществами в регионах за пределами тех, которые традиционно ассоциировались с саладоидным культурным регионом (т.е. Карибский бассейн и нижняя часть долины реки Ориноко). С другой стороны, украшения из нефрита были вырезаны из различных материалов, обычно называемых социальными нефритами, а именно из нефрита, серпентина, хризопраза, альбитита и кварцитов, таких как фуксит. Многие из этих социальных нефритов могли быть добыты на соседних островах. Однако на данный момент источники этих артефактов неизвестны. «Бирюзовые» предметы были идентифицированы как малахит и псевдомалахит с примесью бирюзы. Малахит и бирюза образуются в сухом климате, выветриваются из медных руд и часто связаны с отложениями известняка (Джордж Харлоу, личное сообщение, 2008 г.).Месторождения меди, связанные с дуговым вулканизмом, расположены на Пуэрто-Рико, Виргинских островах (Сент-Джон) и Малых Антильских островах, и Кокс и Бриггс (1973: 3) отметили, что все месторождения меди на Пуэрто-Рико «содержат некоторое количество малахита или азурита. »
»
Резюме
Свидетельства, представленные в этом исследовании геохимического происхождения артефактов, иллюстрируют сложные взаимодействия и потенциальные отношения, которые могут существовать в предполагаемых простых эгалитарных обществах. Стилистические изменения в керамике на Св.Круа и все Виргинские острова указывают на некоторую степень взаимодействия с сообществами в восточной части Пуэрто-Рико и восточной части Эспаньолы. Эти отношения были подтверждены химическими данными, полученными в результате исследования керамики: ранние изделия были импортированы из Пуэрто-Рико, а изделия из карася были доставлены в Пуэрто-Рико и Испаньолу (Доминиканская Республика), что указывает на то, что между этими сообществами существовали некие отношения обмена на большие расстояния. почти 1500 лет назад.
Исследование также показало, что производство практически всей протестированной керамики ограничивалось лишь несколькими местами, расположенными вблизи определенных видов глин.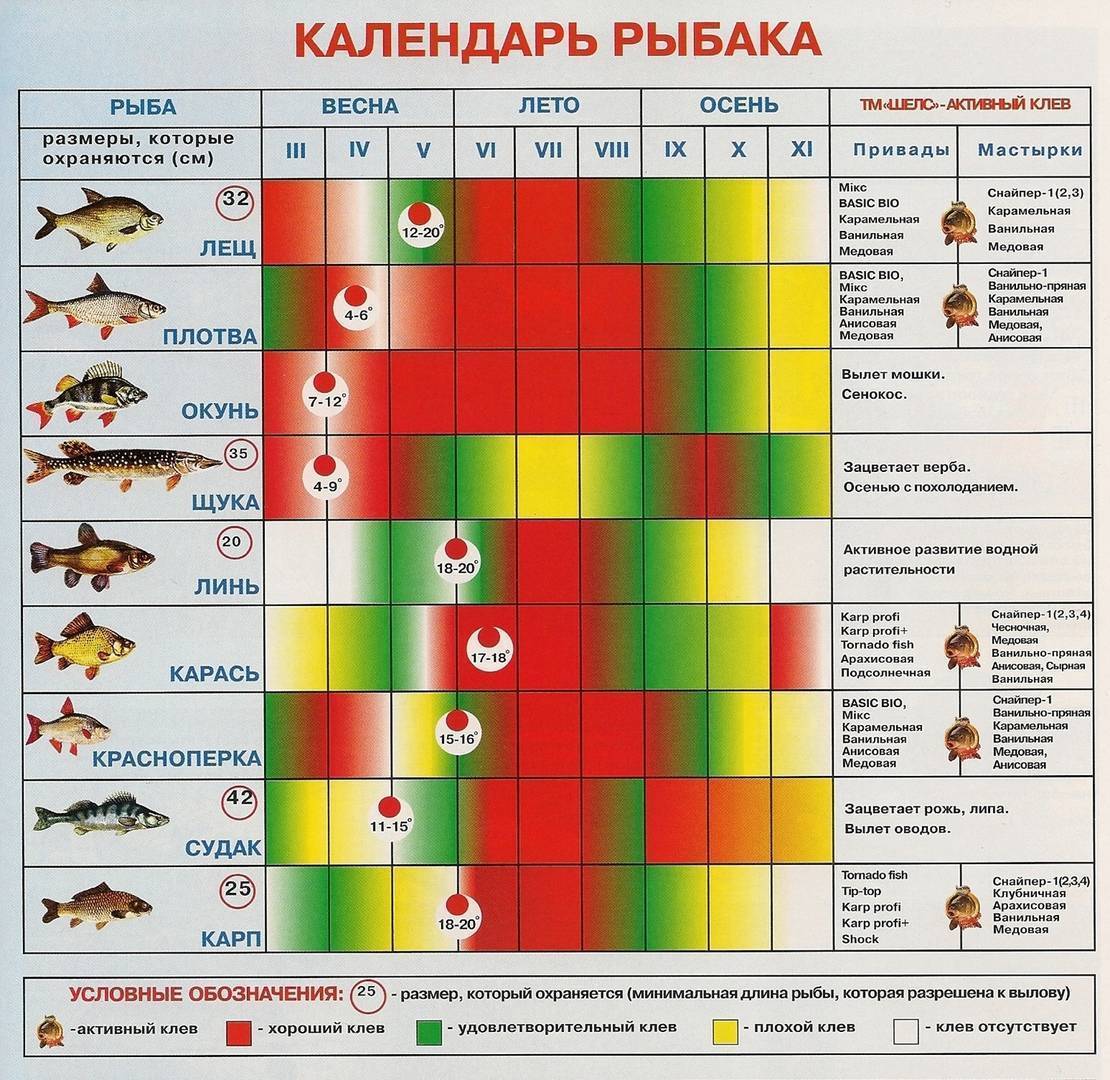 Керамика, или глина, затем перевозилась в другие деревни по всему острову. Основываясь на результатах анализа NAA и сравнениях с другими геохимическими исследованиями на соседних островах, кажется, что гончарное производство было в основном местным и велось на острове. Также не было различий между типами используемых глин и керамическими стилями: изделия WOR и ZIC, изысканные и утилитарные изделия, полые формы и сковороды были сделаны в основном из одних и тех же видов глины.
Керамика, или глина, затем перевозилась в другие деревни по всему острову. Основываясь на результатах анализа NAA и сравнениях с другими геохимическими исследованиями на соседних островах, кажется, что гончарное производство было в основном местным и велось на острове. Также не было различий между типами используемых глин и керамическими стилями: изделия WOR и ZIC, изысканные и утилитарные изделия, полые формы и сковороды были сделаны в основном из одних и тех же видов глины.
Что касается двух видов каменных артефактов, изучаемых здесь, каждый тип был сделан из двух разных камней, которые напоминали друг друга по цвету: настоящий жадеит (кельты) и социальные нефриты, такие как нефрит, серпентин, альбит и хризопраз (украшения).
Таким образом, многие археологические раскопки периода Саладоидов, как на Санта-Крус, так и на других островах северных Малых и Больших Антильских островов, предоставили доказательства того, что их жители участвовали в региональных сферах взаимодействия. Эти свидетельства представлены в виде бус и украшений из камня и ракушек, отшлифованных каменных кельтов из жадеита, добытого из далеких мест, и хорошо сделанной керамики. Некоторые из этих сообществ (возможно, известные как Дома), вероятно, функционировали как специализированные производственные центры, работающие неполный рабочий день.В некоторых случаях эти производственные деревни не обязательно располагались рядом с источниками сырья, что указывало на то, что сырье перемещалось из пункта происхождения в место производства, а затем, возможно, в конечный пункт назначения.
Эти свидетельства представлены в виде бус и украшений из камня и ракушек, отшлифованных каменных кельтов из жадеита, добытого из далеких мест, и хорошо сделанной керамики. Некоторые из этих сообществ (возможно, известные как Дома), вероятно, функционировали как специализированные производственные центры, работающие неполный рабочий день.В некоторых случаях эти производственные деревни не обязательно располагались рядом с источниками сырья, что указывало на то, что сырье перемещалось из пункта происхождения в место производства, а затем, возможно, в конечный пункт назначения.
Это исследование помогло сотрудникам Службы национальных парков в CHRI, SARI и SEAC лучше интерпретировать посетителям парков доисторический образ жизни древних народов Санта-Крус и Виргинских островов. Это позволило улучшить управление и защиту культурного ландшафта и древнего наследия жителей Виргинских островов.Это исследование также продемонстрировало важность и полезность музейных коллекций для интерпретации, образования и научных исследований древнего прошлого.
Благодарности
Особая благодарность бывшим директорам SEAC Бенни Килу и Джону Эренхарду, исполняющему обязанности директора SEAC Джорджу Смиту и Гаю Прентису, менеджеру Региональной программы археологических исследований, за их поддержку и помощь в ходе этого исследования. Нейтронно-активационный анализ был проведен Джеффом Фергюсоном и Майклом Гласкоком из Лаборатории археометрии Исследовательского реактора Университета Миссури, а электронный микрозонд и сканирующий электронный микроскоп были проведены Джорджем Харлоу из Департамента наук о Земле и планетах Американского музея естественной истории.Особая благодарность Биргит Фабер Морс. Наконец, мы выражаем благодарность суперинтенданту Джоэлу Тутейну и руководителю отдела управления ресурсами Зэнди Хиллис-Старр, Национальное историческое место и экологический заповедник Солт-Ривер-Бей и Национальное историческое место Кристианстед, Санта-Крус, за всю их поддержку и интерес к этому исследованию в течение последнего времени. несколько лет. Большая часть этого исследования была оплачена грантом Олбрайт Вирт Службы национальных парков (2005 г.) и Национальным историческим памятником Кристианстед (Служба национальных парков).
Ссылки
Boomert, A.
1987 Дары амазонок: подвески и бусы из зеленого камня как предметы церемониального обмена в Амазонии и Карибском бассейне. Антропологика 67: 33-54.
Карини, С.П.
1991 Композиционный анализ западно-индийской саладоидной керамики и ее отношение к предыстории Пуэрто-Рико. Неопубликованная докторская степень. диссертация, Университет Коннектикута, Сторрс.
Коди, А.
1990 Доисторические модели обмена на Малых Антильских островах: материалы, модели и предварительные наблюдения.Неопубликованная диссертация на степень магистра гуманитарных наук, Государственный университет Сан-Диего.
1991 Распространение каменных артефактов через Малые Антильские острова: их значение для доисторического взаимодействия и обмена. В материалах Четырнадцатого конгресса Международной ассоциации карибской археологии, под редакцией А. Камминса и П. Кинга, стр. 589–604. Барбадосский музей и историческое общество, гарнизон Святой Анны, Сент-Майкл, Барбадос.
Камминса и П. Кинга, стр. 589–604. Барбадосский музей и историческое общество, гарнизон Святой Анны, Сент-Майкл, Барбадос.
Костин, К.Л.
2007 Размышление о производстве: феноменологическая классификация и лексическая семантика.В книге «Переосмысление ремесленной специализации в сложных обществах: археологический анализ социального значения производства», под редакцией Закари X. Хруби и Роуэна К. Флэда, стр. 143–162. Археологические документы Американской антропологической ассоциации, номер 17, Ганновер.
Костин С.Л. и М.Б. Хагструм
1995 Стандартизация, трудовые инвестиции, навыки и организация керамического производства в позднем доиспанском высокогорье Перу. Американская древность 60 (4): 619-639.
Кокс, Д. П.и RP Briggs
1973 Металлогеническая карта Пуэрто-Рико. Разное Карта геологических исследований I-721. Геологическая служба США, Вашингтон, округ Колумбия
Крок, Дж. Г. и Р. Н. Бартоне
1998 г. Археология Трантс, Монтсеррат. Часть 4. Производство чешуйчатого камня и каменных бус. Анналы Музея Карнеги 67 (3): 197-224.
Производство чешуйчатого камня и каменных бус. Анналы Музея Карнеги 67 (3): 197-224.
Фергюсон, Дж. Р. и М. Д. Гласкок
2006 г. Инструментальный нейтронно-активационный анализ Поздняя саладоидная керамика из Санта-Крус, Виргинские острова США. Лаборатория археометрии Исследовательский реактор Университета Миссури Университет Миссури, Колумбия.
Харди, М.
2008 Саладоидная экономика и сложность на границе Аравакана. Неопубликованная докторская степень. диссертация, факультет антропологии, Университет штата Флорида, Таллахасси.
Харлоу, Г.Э., А.Р. Мерфи, Д.Дж. Хозджан, К.Н. де Милль и А.А. Левинсон
2006 г. Доколумбовые жадеитовые топоры из Антигуа, Вест-Индия: описание и возможные источники. Канадский минералог 44 (2): 305-321.
Келкер, Н. Л.
1985 Историческая традиция в искусстве доколумбовой юго-западной Карибской низменности.Неопубликованная докторская степень. диссертация, Техасский университет, Остин.
Knippenberg, S.
2006 г. Производство и обмен каменными артефактами на Северных Малых Антильских островах. Неопубликованная докторская степень. диссертация, кафедра карибской археологии, Лейденский университет, Лейден, Нидерланды.
Производство и обмен каменными артефактами на Северных Малых Антильских островах. Неопубликованная докторская степень. диссертация, кафедра карибской археологии, Лейденский университет, Лейден, Нидерланды.
Мерфи, А. Р., Д. Дж. Хозджан, С. Х. де Милль и А. А. Левинсон
2000 Доколумбовые драгоценные камни и декоративные материалы из Антигуа, Вест-Индия. Драгоценные камни и геммология: 234-245.
МакГиннис, С.А.М. 1997 Идеографическое выражение в доколумбовом Карибском бассейне. Неопубликованная докторская степень. диссертация, Техасский университет, Остин.
Narganes Storde, YM
1995a La Lapidaria de La Hueca, Вьекес, Пуэрто-Рико. В материалах Пятнадцатого конгресса Международной ассоциации карибской археологии, под редакцией Р. Э. Алегрии и М. Родригеса, стр. 141–151. Centro de Estudios Avanzados de Puerto Rico y el Caribe, Сан-Хуан, Пуэрто-Рико.
1995b Lapidaria de Sorce, Vieques, y Tecla, Guayanilla, Пуэрто-Рико.В материалах Шестнадцатого конгресса Международной ассоциации карибской археологии, стр. 17-26. Бас-Тер, Гваделупа.
O’Connor, BC
1999 Поиск закалки и глины для керамики на отдельных участках Виргинских островов Соединенных Штатов. В материалах семнадцатого конгресса Международной ассоциации карибской археологии, под редакцией Дж. Х. Винтера, стр. 148–153. Колледж Моллой, Нью-Йорк.
Поттер, Д.
1996 Химический анализ глины и доисторической керамики из Св.Джон, Виргинские острова США, использование PIXE и рентгеновского дифракционного анализа. Неопубликованная магистерская диссертация, Университет штата Флорида, Таллахасси.
Родригес, М.
1991 Arqueologica de Punta Candelero, Пуэрто-Рико. В материалах Четырнадцатого конгресса Международной ассоциации карибской археологии, под редакцией А. Камминса и П. Кинга, стр. 605–627. Барбадосский музей и историческое общество, гарнизон Святой Анны, Сент-Майкл, Барбадос.
Уоттерс, Д. Р. и Р. Скальон
1994 г. Бусы и подвески из Трантса, Монтсеррат: значение для доисторической гранильной промышленности Карибского бассейна. Анналы музея Карнеги, 63 (3): 215–237.
Анналы музея Карнеги, 63 (3): 215–237.
Загрязнение микропластиком коммерческой соли для потребления человеком: обзор
https://doi.org/10.1016/j.ecss.2019.02.018Get rights and contentHighlights
- •
Пластик как морской мусор — новое дополнение в список глобальных угроз.
- •
Загрязнение моря, несомненно, приведет к загрязнению морепродуктов.
- •
Микропластик в солях может представлять угрозу безопасности пищевых продуктов и здоровью людей.
- •
Микропластик поглощает загрязняющие вещества и переносит их в соль и другие продукты.
Abstract
Микропластики (МП) представляют собой пластиковые частицы размером менее 5 мм, которые считаются глобальными загрязнителями окружающей среды. Присутствующие в окружающей среде МЧ возникают в результате последовательного распада более крупных кусков пластика или прямого поступления микро- и наночастиц, используемых в различных отраслях промышленности и в продуктах, доступных потребителям. Такие MP были обнаружены в нескольких диких видах и других природных ресурсах, в том числе в некоторых из них, потребляемых людьми в пищу, с возможным неблагоприятным воздействием на экосистему и здоровье человека. Основная цель этой работы заключалась в обзоре опубликованной литературы о загрязнении морских коммерческих солей (морского и наземного происхождения) и его возможном воздействии на здоровье человека. Кроме того, чтобы привести к всестороннему пониманию парадигмы, включено краткое введение и пересмотр загрязнения окружающей среды MP и его последствий.MP были обнаружены в коммерческих солях 128 брендов из 38 разных стран на пяти континентах. Концентрация МЧ, обнаруженная в проанализированных образцах, ниже, чем концентрация МЧ, зарегистрированная в других ресурсах, таких как голубые мидии. Однако, поскольку коммерческие соли используются каждый день и всеми людьми, они представляют собой путь долгосрочного воздействия на население в целом в дополнение к другим (например, животные, употребляемые в пищу людьми, вода, воздух). Таким образом, коммерческие соли, загрязненные МП, могут способствовать потенциальному долгосрочному неблагоприятному воздействию в результате воздействия этих частиц на человека.
Такие MP были обнаружены в нескольких диких видах и других природных ресурсах, в том числе в некоторых из них, потребляемых людьми в пищу, с возможным неблагоприятным воздействием на экосистему и здоровье человека. Основная цель этой работы заключалась в обзоре опубликованной литературы о загрязнении морских коммерческих солей (морского и наземного происхождения) и его возможном воздействии на здоровье человека. Кроме того, чтобы привести к всестороннему пониманию парадигмы, включено краткое введение и пересмотр загрязнения окружающей среды MP и его последствий.MP были обнаружены в коммерческих солях 128 брендов из 38 разных стран на пяти континентах. Концентрация МЧ, обнаруженная в проанализированных образцах, ниже, чем концентрация МЧ, зарегистрированная в других ресурсах, таких как голубые мидии. Однако, поскольку коммерческие соли используются каждый день и всеми людьми, они представляют собой путь долгосрочного воздействия на население в целом в дополнение к другим (например, животные, употребляемые в пищу людьми, вода, воздух). Таким образом, коммерческие соли, загрязненные МП, могут способствовать потенциальному долгосрочному неблагоприятному воздействию в результате воздействия этих частиц на человека.
Ключевые слова
Микропластик
Глобальные загрязнители окружающей среды
Воздействие на окружающую среду
Коммерческие соли для потребления человеком
Безопасность пищевых продуктов и здоровье человека
Рекомендуемые статьиСсылки на статьи (0)
Опубликовано Elsevier Ltd.
Рекомендуемые статьи
Ссылки на статьи
Animal Crossing: New Leaf События июля | Стратегия
Animal Crossing: New Leaf — это игра, в которой, если вы не обратите внимание, вы обязательно пропустите что-то интересное.Оказывается, каждый месяц вы получаете совершенно новый контент, будь то особый праздник страны, эксклюзивная рыба или чей-то день рождения.
Июль не исключение, и для вашего удобства мы перечислили все праздники, доступную рыбу, насекомых и глубоководный улов. Мы также разблокировали эту информацию из нашего официального Animal Crossing: New Leaf Guide , который вы можете приобрести за 9,99 долларов США.
Мы также разблокировали эту информацию из нашего официального Animal Crossing: New Leaf Guide , который вы можете приобрести за 9,99 долларов США.
Особые события
День звездных крестов (Япония): посетите друга в Японии и насладитесь празднованием дня.
Bug-Off (3-я суббота): возьмите свою лучшую сеть и посмотрите, сможете ли вы поймать жука дня!
День рождения – будь милым и дари подарки
1 июля: День рождения Курта «Пушистый комочек»
2 июля: День рождения агента S, «Напарник»
3 июля: День рождения Блэр «Орешек»
4 июля: День рождения Аполлона «Фа»
5 июля: День рождения Самсона «Крошка»
6 июля: День рождения Миры «Хвост»
7 июля: День рождения Бри «Чизбол»
8 июля: День рождения Иеремии «Ни-дип»
9 июля: День рождения Статика «Крззт»
10 июля: День рождения Мотта «Кейджи»
11 июля: День рождения Виктории «Сахарный кубик»
12 июля: нет
13 июля: День рождения Твигги «Чиперс»
14 июля: День рождения Диззи «Ву-у-у»
15 июля: День рождения Кэролайн «Хулаааа»
16 июля: День рождения Фриты «О, овца»
17 июля: День рождения Джея «Хееееей»
18 июля: День Рождения Крока «Как будто»
19 июля: День рождения Ниббла «Ниббл»
20 июля: День рождения Юки «Тск-тск»
21 июля: День рождения принца «Бурруп»
22 июля: День рождения Чоу «Айя»
23 июля: День рождения Элвиса «Угу»
24 июля: День рождения О’Хара «Амиго»
25 июля: День рождения Пека «Хруст»
26 июля: День рождения Керли «Нёинк»
27 июля: День рождения Эрика «Поужинайте»
28 июля: День рождения трюфеля «Морда»
29 июля: День рождения Лайонела «Точно»
30 июля: День рождения Фрэнка «Crushy»
31 июля: День рождения Гризли «Гррр…»
Доступные насекомые: Бабочка Агриас, Муравей, Рогатый атлас, Полосатая стрекоза, Пчела, Птицекрыл, Коричневая цикада, Многоножка, Панцирь цикады, Цикломатус, Стрекоза Дарнер, Плавучий жук, Рогатый династид, Рогатый слон, Императорская бабочка, Вечерняя цикада, Блоха , Муха, Фруктовый жук, Гигантский олень, Гигантская цикада, Золотой олень. Жук-голиаф, Кузнечик, Рогатый Геркулес, Краб-отшельник, Медоносная пчела, Жук-драгоценность, Фонарная муха, Саранча, Длиннорогий жук, Богомол, Олень Мияма, Комар, Мотылек, Дубовый шелковый мотылек, Орхидейный богомол, Бабочка-павлин, Жук-таблетка, Прудоскатер, Радуга Олень, Раджа Брук, Крепкая цикада, Олень-пила, Жук-скарабей, Скорпион, Улитка, Паук, Жук-вонючка, Тарантул, Жук-тигр, Бабочка-тигр, Цикада-ходок, Ходячий лист, Трость, Причаловая плотва.
Жук-голиаф, Кузнечик, Рогатый Геркулес, Краб-отшельник, Медоносная пчела, Жук-драгоценность, Фонарная муха, Саранча, Длиннорогий жук, Богомол, Олень Мияма, Комар, Мотылек, Дубовый шелковый мотылек, Орхидейный богомол, Бабочка-павлин, Жук-таблетка, Прудоскатер, Радуга Олень, Раджа Брук, Крепкая цикада, Олень-пила, Жук-скарабей, Скорпион, Улитка, Паук, Жук-вонючка, Тарантул, Жук-тигр, Бабочка-тигр, Цикада-ходок, Ходячий лист, Трость, Причаловая плотва.
Доступная рыба: Рыба-ангел, арапайма, арована, усач, полосатый нож, черный окунь, голубой марлин, голубогиль, рыба-бабочка, карп, сом, рыба-клоун, латимерия, раки, карась, елец, дорадо, угорь, пресноводный бычок , Лягушка, Гар, Гигантский змееголов, Гигантский Тревалли, Золотая рыбка, Гуппи, Акула-молот, Скумбрия, Киллифиш, Кои, Рыба-наполеон, Неоновая тетра, Рыба-клев, Морская солнечная рыба, Оливковая камбала, Бледный голавль, Пиранья, Золотая рыбка с попоглазыми глазами, Рыба-фугу, Красный люциан, полосатый угорь, оседланный бичир, акула-пила, морской окунь, морской конек, акула, кальмар, рыба-хирург, сладкая рыба, головастик, китовая акула, индейка-зебра.
Доступные глубоководные уловы: морское ушко, желудевый ракушка, наутилус с камерой, ушная раковина, гигантский изопод, мечехвост, омар, креветка-богомол, осьминог, жемчужная устрица, гребешок, актиния, морской виноград, морской слизень, морская звезда, Морской еж, водоросли, пятнистый садовый угорь, тигровая креветка.
Animal Crossing: New Leaf доступен как в розничном, так и в цифровом формате для Nintendo 3DS.
Руководство Prima Games по игре Animal Crossing: New Leaf уже доступно. Разблокируйте его здесь .
Дополнительные функции Animal Crossing
Советы начинающим
Электронный гид по острову Тортимер
Насадки для рыбалки
Советы по отлову насекомых
Обучение лицензированных ветеринаров-рыболовов борьбе с остатками наркотиков в аквакультуре в провинции Гуандун на JSTOR
Абстрактный Китай является крупнейшим в мире производителем аквакультуры, на долю которого приходится 63% мирового производства по объему. Однако с 2000-х годов репутация китайских морепродуктов была испорчена серией инцидентов с остатками наркотиков. Необходимость обеспечения безопасности пищевых продуктов в сочетании с решимостью государства выполнить свои обязательства по борьбе с эпидемиями среди животных после атипичной пневмонии (тяжелого острого респираторного синдрома) и птичьего гриппа вынудили правительство начать реформу ветеринарной системы. Эта реформа была направлена на преобразование квазигосударственных сельских ветеринаров в чувствительных к рынку и технически подкованных лицензированных специалистов, которые могут дисциплинировать непослушных рыбоводов.Тем не менее, ветеринары столкнулись со многими дилеммами, пытаясь найти баланс между безопасностью пищевых продуктов и прибылью от лекарств, здоровьем животных и здоровьем человека, нормативными требованиями и практическими потребностями самих фермеров. В статье показано, как неолиберальная программа воспитания рациональных и ответственных ветеринаров скорее перераспределяет, чем минимизирует риски, возникающие в результате ориентированного на рынок производства аквакультуры.
Однако с 2000-х годов репутация китайских морепродуктов была испорчена серией инцидентов с остатками наркотиков. Необходимость обеспечения безопасности пищевых продуктов в сочетании с решимостью государства выполнить свои обязательства по борьбе с эпидемиями среди животных после атипичной пневмонии (тяжелого острого респираторного синдрома) и птичьего гриппа вынудили правительство начать реформу ветеринарной системы. Эта реформа была направлена на преобразование квазигосударственных сельских ветеринаров в чувствительных к рынку и технически подкованных лицензированных специалистов, которые могут дисциплинировать непослушных рыбоводов.Тем не менее, ветеринары столкнулись со многими дилеммами, пытаясь найти баланс между безопасностью пищевых продуктов и прибылью от лекарств, здоровьем животных и здоровьем человека, нормативными требованиями и практическими потребностями самих фермеров. В статье показано, как неолиберальная программа воспитания рациональных и ответственных ветеринаров скорее перераспределяет, чем минимизирует риски, возникающие в результате ориентированного на рынок производства аквакультуры.
Охватывая весь спектр китаеведения за шесть столетий, «Современный Китай» поощряет науку, которая пересекает старые границы «досовременный/современный» и «современный/современный».В дополнение к исследовательским статьям «Современный Китай» представляет периодические симпозиумы по важным темам китаеведения, критические очерки о состоянии китаеведения, подробные обзорные статьи по конкретным областям науки и обзоры книг необычного качества и значимости.
Информация об издателе Сара Миллер МакКьюн основала издательство SAGE Publishing в 1965 году для поддержки распространения полезных знаний и просвещения мирового сообщества. SAGE является ведущим международным поставщиком инновационного высококачественного контента, который ежегодно публикует более 900 журналов и более 800 новых книг, охватывающих широкий спектр предметных областей.Растущий выбор библиотечных продуктов включает архивы, данные, тематические исследования и видео. Контрольный пакет SAGE по-прежнему принадлежит нашему основателю, а после ее жизни перейдет в собственность благотворительного фонда, который обеспечивает постоянную независимость компании. Основные офисы расположены в Лос-Анджелесе, Лондоне, Нью-Дели, Сингапуре, Вашингтоне и Мельбурне. www.sagepublishing.com
Контрольный пакет SAGE по-прежнему принадлежит нашему основателю, а после ее жизни перейдет в собственность благотворительного фонда, который обеспечивает постоянную независимость компании. Основные офисы расположены в Лос-Анджелесе, Лондоне, Нью-Дели, Сингапуре, Вашингтоне и Мельбурне. www.sagepublishing.com
Конгресс.гов | Библиотека Конгресса
Раздел протокола Конгресса Ежедневный дайджест Сенат дом Расширения замечаний
Замечания участников
Автор Any House MemberАдамс, Альма С. [D-NC] Адерхольт, Роберт Б. [R-AL] Агилар, Пит [D-CA] Аллен, Рик В. [R-GA] Оллред, Колин З. [D-TX] Амодеи, Марк Э. [R -NV] Армстронг, Келли [R-ND] Аррингтон, Джоди С. [R-TX] Окинклосс, Джейк [D-MA] Эксн, Синтия [D-IA] Бабин, Брайан [R-TX] Бэкон, Дон [R -NE] Бэрд, Джеймс Р. [R-IN] Балдерсон, Трой [R-OH] Бэнкс, Джим [R-IN] Барр, Энди [R-KY] Барраган, Нанетт Диаз [D-CA] Басс, Карен [ D-CA] Битти, Джойс [D-OH] Бенц, Клифф [R-OR] Бера, Ами [D-CA] Бергман, Джек [R-MI] Бейер, Дональд С.-младший [D-VA] Байс , Стефани И. [R-OK] Биггс, Энди [R-AZ] Билиракис, Гас М.[R-FL] Бишоп, Дэн [R-NC] Бишоп, Сэнфорд Д., младший [D-GA] Блюменауэр, Эрл [D-OR] Блант Рочестер, Лиза [D-DE] Боберт, Лорен [R-CO ] Бонамичи, Сюзанна [D-OR] Бост, Майк [R-IL] Бурдо, Кэролайн [D-GA] Боуман, Джамаал [D-NY] Бойл, Брендан Ф. [D-PA] Брэди, Кевин [R-TX ] Брукс, Мо [R-AL] Браун, Энтони Г. [D-MD] Браун, Шонтел М. [D-OH] Браунли, Джулия [D-CA] Бьюкенен, Верн [R-FL] Бак, Кен [R -CO] Бакшон, Ларри [R-IN] Бадд, Тед [R-NC] Берчетт, Тим [R-TN] Берджесс, Майкл С.
[D-NC] Адерхольт, Роберт Б. [R-AL] Агилар, Пит [D-CA] Аллен, Рик В. [R-GA] Оллред, Колин З. [D-TX] Амодеи, Марк Э. [R -NV] Армстронг, Келли [R-ND] Аррингтон, Джоди С. [R-TX] Окинклосс, Джейк [D-MA] Эксн, Синтия [D-IA] Бабин, Брайан [R-TX] Бэкон, Дон [R -NE] Бэрд, Джеймс Р. [R-IN] Балдерсон, Трой [R-OH] Бэнкс, Джим [R-IN] Барр, Энди [R-KY] Барраган, Нанетт Диаз [D-CA] Басс, Карен [ D-CA] Битти, Джойс [D-OH] Бенц, Клифф [R-OR] Бера, Ами [D-CA] Бергман, Джек [R-MI] Бейер, Дональд С.-младший [D-VA] Байс , Стефани И. [R-OK] Биггс, Энди [R-AZ] Билиракис, Гас М.[R-FL] Бишоп, Дэн [R-NC] Бишоп, Сэнфорд Д., младший [D-GA] Блюменауэр, Эрл [D-OR] Блант Рочестер, Лиза [D-DE] Боберт, Лорен [R-CO ] Бонамичи, Сюзанна [D-OR] Бост, Майк [R-IL] Бурдо, Кэролайн [D-GA] Боуман, Джамаал [D-NY] Бойл, Брендан Ф. [D-PA] Брэди, Кевин [R-TX ] Брукс, Мо [R-AL] Браун, Энтони Г. [D-MD] Браун, Шонтел М. [D-OH] Браунли, Джулия [D-CA] Бьюкенен, Верн [R-FL] Бак, Кен [R -CO] Бакшон, Ларри [R-IN] Бадд, Тед [R-NC] Берчетт, Тим [R-TN] Берджесс, Майкл С. [R-TX] Буш, Кори [D-MO] Бустос, Чери [D -ИЛ] Баттерфилд, Г.К. [D-NC] Калверт, Кен [R-CA] Каммак, Кэт [R-FL] Карбахал, Салуд О. [D-CA] Карденас, Тони [D-CA] Кэри, Майк [R-OH] Карл , Джерри Л. [R-AL] Карсон, Андре [D-IN] Картер, Эрл Л. «Бадди» [R-GA] Картер, Джон Р. [R-TX] Картер, Трой [D-LA] Картрайт, Мэтт [D-PA] Кейс, Эд [D-HI] Кастен, Шон [D-IL] Кастор, Кэти [D-FL] Кастро, Хоакин [D-TX] Коуторн, Мэдисон [R-NC] Шабо, Стив [ R-OH] Чейни, Лиз [R-WY] Чу, Джуди [D-CA] Чичиллин, Дэвид Н. [D-RI] Кларк, Кэтрин М. [D-MA] Кларк, Иветт Д. [D-NY] Кливер, Эмануэль [D-MO] Клайн, Бен [R-VA] Клауд, Майкл [R-TX] Клайберн, Джеймс Э.[D-SC] Клайд, Эндрю С. [R-GA] Коэн, Стив [D-TN] Коул, Том [R-OK] Комер, Джеймс [R-KY] Коннолли, Джеральд Э. [D-VA] Купер , Джим [D-TN] Корреа, Дж. Луис [D-CA] Коста, Джим [D-CA] Кортни, Джо [D-CT] Крейг, Энджи [D-MN] Кроуфорд, Эрик А. «Рик» [ R-AR] Креншоу, Дэн [R-TX] Крист, Чарли [D-FL] Кроу, Джейсон [D-CO] Куэльяр, Генри [D-TX] Кертис, Джон Р. [R-UT] Дэвидс, Шарис [ D-KS] Дэвидсон, Уоррен [R-OH] Дэвис, Дэнни К.
[R-TX] Буш, Кори [D-MO] Бустос, Чери [D -ИЛ] Баттерфилд, Г.К. [D-NC] Калверт, Кен [R-CA] Каммак, Кэт [R-FL] Карбахал, Салуд О. [D-CA] Карденас, Тони [D-CA] Кэри, Майк [R-OH] Карл , Джерри Л. [R-AL] Карсон, Андре [D-IN] Картер, Эрл Л. «Бадди» [R-GA] Картер, Джон Р. [R-TX] Картер, Трой [D-LA] Картрайт, Мэтт [D-PA] Кейс, Эд [D-HI] Кастен, Шон [D-IL] Кастор, Кэти [D-FL] Кастро, Хоакин [D-TX] Коуторн, Мэдисон [R-NC] Шабо, Стив [ R-OH] Чейни, Лиз [R-WY] Чу, Джуди [D-CA] Чичиллин, Дэвид Н. [D-RI] Кларк, Кэтрин М. [D-MA] Кларк, Иветт Д. [D-NY] Кливер, Эмануэль [D-MO] Клайн, Бен [R-VA] Клауд, Майкл [R-TX] Клайберн, Джеймс Э.[D-SC] Клайд, Эндрю С. [R-GA] Коэн, Стив [D-TN] Коул, Том [R-OK] Комер, Джеймс [R-KY] Коннолли, Джеральд Э. [D-VA] Купер , Джим [D-TN] Корреа, Дж. Луис [D-CA] Коста, Джим [D-CA] Кортни, Джо [D-CT] Крейг, Энджи [D-MN] Кроуфорд, Эрик А. «Рик» [ R-AR] Креншоу, Дэн [R-TX] Крист, Чарли [D-FL] Кроу, Джейсон [D-CO] Куэльяр, Генри [D-TX] Кертис, Джон Р. [R-UT] Дэвидс, Шарис [ D-KS] Дэвидсон, Уоррен [R-OH] Дэвис, Дэнни К. [D-IL] Дэвис, Родни [R-IL] Дин, Мадлен [D-PA] ДеФацио, Питер А. [D-OR] ДеГетт, Диана [D-CO] ДеЛауро, Роза Л.[D-CT] ДельБен, Сьюзан К. [D-WA] Дельгадо, Антонио [D-NY] Демингс, Вэл Батлер [D-FL] ДеСольнье, Марк [D-CA] Дежарле, Скотт [R-TN] Дойч, Теодор Э. [D-FL] Диас-Баларт, Марио [R-FL] Дингелл, Дебби [D-MI] Доггетт, Ллойд [D-TX] Дональдс, Байрон [R-FL] Дойл, Майкл Ф. [D- PA] Дункан, Джефф [R-SC] Данн, Нил П. [R-FL] Эллзи, Джейк [R-TX] Эммер, Том [R-MN] Эскобар, Вероника [D-TX] Эшу, Анна Г. [ D-CA] Эспайлат, Адриано [D-NY] Эстес, Рон [R-KS] Эванс, Дуайт [D-PA] Фаллон, Пэт [R-TX] Финстра, Рэнди [R-IA] Фергюсон, А. Дрю, IV [R-GA] Фишбах, Мишель [R-MN] Фицджеральд, Скотт [R-WI] Фицпатрик, Брайан К.[R-PA] Флейшманн, Чарльз Дж. «Чак» [R-TN] Флетчер, Лиззи [D-TX] Фортенберри, Джефф [R-NE] Фостер, Билл [D-IL] Фокс, Вирджиния [R-NC] Франкель, Лоис [D-FL] Франклин, К. Скотт [R-FL] Фадж, Марсия Л. [D-OH] Фулчер, Расс [R-ID] Гэтц, Мэтт [R-FL] Галлахер, Майк [R- WI] Гальего, Рубен [D-AZ] Гараменди, Джон [D-CA] Гарбарино, Эндрю Р. [R-NY] Гарсия, Хесус Г. «Чуй» [D-IL] Гарсия, Майк [R-CA] Гарсия , Сильвия Р. [D-TX] Гиббс, Боб [R-OH] Гименес, Карлос А. [R-FL] Гомерт, Луи [R-TX] Голден, Джаред Ф. [D-ME] Гомес, Джимми [D -CA] Гонсалес, Тони [R-TX] Гонсалес, Энтони [R-OH] Гонсалес, Висенте [D-TX] Гонсалес-Колон, Дженниффер [R-PR] Гуд, Боб [R-VA] Гуден, Лэнс [R -TX] Госар, Пол А.[R-AZ] Готхаймер, Джош [D-NJ] Грейнджер, Кей [R-TX] Грейвс, Гаррет [R-LA] Грейвс, Сэм [R-MO] Грин, Эл [D-TX] Грин, Марк Э. [R-TN] Грин, Марджори Тейлор [R-GA] Гриффит, Х. Морган [R-VA] Грихальва, Рауль М. [D-AZ] Гротман, Гленн [R-WI] Гест, Майкл [R-MS] Гатри, Бретт [R-KY] Хааланд, Дебра А. [D-NM] Хагедорн, Джим [R-MN] Хардер, Джош [D-CA] Харрис, Энди [R-MD] Харшбаргер, Диана [R-TN] Хартцлер, Вики [R-MO] Гастингс, Элси Л. [D-FL] Хейс, Джахана [D-CT] Херн, Кевин [R-OK] Херрелл, Иветт [R-NM] Эррера Бейтлер, Хайме [R-WA ] Хайс, Джоди Б.[R-GA] Хиггинс, Брайан [D-NY] Хиггинс, Клэй [R-LA] Хилл, Дж. Френч [R-AR] Хаймс, Джеймс А. [D-CT] Хинсон, Эшли [R-IA] Холлингсворт , Трей [R-IN] Хорсфорд, Стивен [D-NV] Хулахан, Крисси [D-PA] Хойер, Стени Х.
[D-IL] Дэвис, Родни [R-IL] Дин, Мадлен [D-PA] ДеФацио, Питер А. [D-OR] ДеГетт, Диана [D-CO] ДеЛауро, Роза Л.[D-CT] ДельБен, Сьюзан К. [D-WA] Дельгадо, Антонио [D-NY] Демингс, Вэл Батлер [D-FL] ДеСольнье, Марк [D-CA] Дежарле, Скотт [R-TN] Дойч, Теодор Э. [D-FL] Диас-Баларт, Марио [R-FL] Дингелл, Дебби [D-MI] Доггетт, Ллойд [D-TX] Дональдс, Байрон [R-FL] Дойл, Майкл Ф. [D- PA] Дункан, Джефф [R-SC] Данн, Нил П. [R-FL] Эллзи, Джейк [R-TX] Эммер, Том [R-MN] Эскобар, Вероника [D-TX] Эшу, Анна Г. [ D-CA] Эспайлат, Адриано [D-NY] Эстес, Рон [R-KS] Эванс, Дуайт [D-PA] Фаллон, Пэт [R-TX] Финстра, Рэнди [R-IA] Фергюсон, А. Дрю, IV [R-GA] Фишбах, Мишель [R-MN] Фицджеральд, Скотт [R-WI] Фицпатрик, Брайан К.[R-PA] Флейшманн, Чарльз Дж. «Чак» [R-TN] Флетчер, Лиззи [D-TX] Фортенберри, Джефф [R-NE] Фостер, Билл [D-IL] Фокс, Вирджиния [R-NC] Франкель, Лоис [D-FL] Франклин, К. Скотт [R-FL] Фадж, Марсия Л. [D-OH] Фулчер, Расс [R-ID] Гэтц, Мэтт [R-FL] Галлахер, Майк [R- WI] Гальего, Рубен [D-AZ] Гараменди, Джон [D-CA] Гарбарино, Эндрю Р. [R-NY] Гарсия, Хесус Г. «Чуй» [D-IL] Гарсия, Майк [R-CA] Гарсия , Сильвия Р. [D-TX] Гиббс, Боб [R-OH] Гименес, Карлос А. [R-FL] Гомерт, Луи [R-TX] Голден, Джаред Ф. [D-ME] Гомес, Джимми [D -CA] Гонсалес, Тони [R-TX] Гонсалес, Энтони [R-OH] Гонсалес, Висенте [D-TX] Гонсалес-Колон, Дженниффер [R-PR] Гуд, Боб [R-VA] Гуден, Лэнс [R -TX] Госар, Пол А.[R-AZ] Готхаймер, Джош [D-NJ] Грейнджер, Кей [R-TX] Грейвс, Гаррет [R-LA] Грейвс, Сэм [R-MO] Грин, Эл [D-TX] Грин, Марк Э. [R-TN] Грин, Марджори Тейлор [R-GA] Гриффит, Х. Морган [R-VA] Грихальва, Рауль М. [D-AZ] Гротман, Гленн [R-WI] Гест, Майкл [R-MS] Гатри, Бретт [R-KY] Хааланд, Дебра А. [D-NM] Хагедорн, Джим [R-MN] Хардер, Джош [D-CA] Харрис, Энди [R-MD] Харшбаргер, Диана [R-TN] Хартцлер, Вики [R-MO] Гастингс, Элси Л. [D-FL] Хейс, Джахана [D-CT] Херн, Кевин [R-OK] Херрелл, Иветт [R-NM] Эррера Бейтлер, Хайме [R-WA ] Хайс, Джоди Б.[R-GA] Хиггинс, Брайан [D-NY] Хиггинс, Клэй [R-LA] Хилл, Дж. Френч [R-AR] Хаймс, Джеймс А. [D-CT] Хинсон, Эшли [R-IA] Холлингсворт , Трей [R-IN] Хорсфорд, Стивен [D-NV] Хулахан, Крисси [D-PA] Хойер, Стени Х. [D-MD] Хадсон, Ричард [R-NC] Хаффман, Джаред [D-CA] Хьюзенга , Билл [R-MI] Исса, Даррелл Э. [R-CA] Джексон Ли, Шейла [D-TX] Джексон, Ронни [R-TX] Джейкобс, Крис [R-NY] Джейкобс, Сара [D-CA] Джаяпал, Прамила [D-WA] Джеффрис, Хаким С. [D-NY] Джонсон, Билл [R-OH] Джонсон, Дасти [R-SD] Джонсон, Эдди Бернис [D-TX] Джонсон, Генри С.«Хэнк» младший [D-GA] Джонсон, Майк [R-LA] Джонс, Мондэр [D-NY] Джордан, Джим [R-OH] Джойс, Дэвид П. [R-OH] Джойс, Джон [R -PA] Кахеле, Кайалии [D-HI] Каптур, Марси [D-OH] Катко, Джон [R-NY] Китинг, Уильям Р. [D-MA] Келлер, Фред [R-PA] Келли, Майк [R-PA] Келли, Робин Л. [D-IL] Келли, Трент [R-MS] Ханна, Ро [D-CA] Килди, Дэниел Т. [D-MI] Килмер, Дерек [D-WA] Ким , Энди [D-NJ] Ким, Янг [R-CA] Кайнд, Рон [D-WI] Кинзингер, Адам [R-IL] Киркпатрик, Энн [D-AZ] Кришнамурти, Раджа [D-IL] Кастер, Энн М. [D-NH] Кастофф, Дэвид [R-TN] ЛаХуд, Дарин [R-IL] ЛаМальфа, Дуг [R-CA] Лэмб, Конор [D-PA] Ламборн, Дуг [R-CO] Ланжевен, Джеймс Р.[D-RI] Ларсен, Рик [D-WA] Ларсон, Джон Б. [D-CT] Латта, Роберт Э.
[D-MD] Хадсон, Ричард [R-NC] Хаффман, Джаред [D-CA] Хьюзенга , Билл [R-MI] Исса, Даррелл Э. [R-CA] Джексон Ли, Шейла [D-TX] Джексон, Ронни [R-TX] Джейкобс, Крис [R-NY] Джейкобс, Сара [D-CA] Джаяпал, Прамила [D-WA] Джеффрис, Хаким С. [D-NY] Джонсон, Билл [R-OH] Джонсон, Дасти [R-SD] Джонсон, Эдди Бернис [D-TX] Джонсон, Генри С.«Хэнк» младший [D-GA] Джонсон, Майк [R-LA] Джонс, Мондэр [D-NY] Джордан, Джим [R-OH] Джойс, Дэвид П. [R-OH] Джойс, Джон [R -PA] Кахеле, Кайалии [D-HI] Каптур, Марси [D-OH] Катко, Джон [R-NY] Китинг, Уильям Р. [D-MA] Келлер, Фред [R-PA] Келли, Майк [R-PA] Келли, Робин Л. [D-IL] Келли, Трент [R-MS] Ханна, Ро [D-CA] Килди, Дэниел Т. [D-MI] Килмер, Дерек [D-WA] Ким , Энди [D-NJ] Ким, Янг [R-CA] Кайнд, Рон [D-WI] Кинзингер, Адам [R-IL] Киркпатрик, Энн [D-AZ] Кришнамурти, Раджа [D-IL] Кастер, Энн М. [D-NH] Кастофф, Дэвид [R-TN] ЛаХуд, Дарин [R-IL] ЛаМальфа, Дуг [R-CA] Лэмб, Конор [D-PA] Ламборн, Дуг [R-CO] Ланжевен, Джеймс Р.[D-RI] Ларсен, Рик [D-WA] Ларсон, Джон Б. [D-CT] Латта, Роберт Э. [R-OH] ЛаТернер, Джейк [R-KS] Лоуренс, Бренда Л. [D-MI ] Лоусон, Эл-младший [D-FL] Ли, Барбара [D-CA] Ли, Сьюзи [D-NV] Леже Фернандес, Тереза [D-NM] Леско, Дебби [R-AZ] Летлоу, Джулия [R -LA] Левин, Энди [D-MI] Левин, Майк [D-CA] Лью, Тед [D-CA] Лофгрен, Зои [D-CA] Лонг, Билли [R-MO] Лоудермилк, Барри [R-GA ]Ловенталь, Алан С. [D-CA]Лукас, Фрэнк Д. [R-OK]Люткемейер, Блейн [R-MO]Лурия, Элейн Г. [D-VA]Линч, Стивен Ф. [D-MA]Мейс , Нэнси [R-SC] Малиновски, Том [D-NJ] Маллиотакис, Николь [R-NY] Мэлони, Кэролин Б.[D-NY] Мэлони, Шон Патрик [D-NY] Манн, Трейси [R-KS] Мэннинг, Кэти Э. [D-NC] Мэсси, Томас [R-KY] Маст, Брайан Дж. [R-FL] Мацуи, Дорис О. [D-CA] МакБат, Люси [D-GA] Маккарти, Кевин [R-CA] МакКол, Майкл Т. [R-TX] Макклейн, Лиза К. [R-MI] МакКлинток, Том [ R-CA] Макколлум, Бетти [D-MN] МакИчин, А. Дональд [D-VA] Макговерн, Джеймс П. [D-MA] МакГенри, Патрик Т. [R-NC] МакКинли, Дэвид Б. [R- WV] Макморрис Роджерс, Кэти [R-WA] Макнерни, Джерри [D-CA] Микс, Грегори В. [D-NY] Мейер, Питер [R-MI] Менг, Грейс [D-NY] Мейзер, Дэниел [R -PA] Мфуме, Квейси [D-MD] Миллер, Кэрол Д.
[R-OH] ЛаТернер, Джейк [R-KS] Лоуренс, Бренда Л. [D-MI ] Лоусон, Эл-младший [D-FL] Ли, Барбара [D-CA] Ли, Сьюзи [D-NV] Леже Фернандес, Тереза [D-NM] Леско, Дебби [R-AZ] Летлоу, Джулия [R -LA] Левин, Энди [D-MI] Левин, Майк [D-CA] Лью, Тед [D-CA] Лофгрен, Зои [D-CA] Лонг, Билли [R-MO] Лоудермилк, Барри [R-GA ]Ловенталь, Алан С. [D-CA]Лукас, Фрэнк Д. [R-OK]Люткемейер, Блейн [R-MO]Лурия, Элейн Г. [D-VA]Линч, Стивен Ф. [D-MA]Мейс , Нэнси [R-SC] Малиновски, Том [D-NJ] Маллиотакис, Николь [R-NY] Мэлони, Кэролин Б.[D-NY] Мэлони, Шон Патрик [D-NY] Манн, Трейси [R-KS] Мэннинг, Кэти Э. [D-NC] Мэсси, Томас [R-KY] Маст, Брайан Дж. [R-FL] Мацуи, Дорис О. [D-CA] МакБат, Люси [D-GA] Маккарти, Кевин [R-CA] МакКол, Майкл Т. [R-TX] Макклейн, Лиза К. [R-MI] МакКлинток, Том [ R-CA] Макколлум, Бетти [D-MN] МакИчин, А. Дональд [D-VA] Макговерн, Джеймс П. [D-MA] МакГенри, Патрик Т. [R-NC] МакКинли, Дэвид Б. [R- WV] Макморрис Роджерс, Кэти [R-WA] Макнерни, Джерри [D-CA] Микс, Грегори В. [D-NY] Мейер, Питер [R-MI] Менг, Грейс [D-NY] Мейзер, Дэниел [R -PA] Мфуме, Квейси [D-MD] Миллер, Кэрол Д. [R-WV] Миллер, Мэри Э. [R-IL] Миллер-Микс, Марианнетт [R-IA] Муленаар, Джон Р. [R-MI] Муни, Александр X. [R-WV] Мур, Барри [R -AL] Мур, Блейк Д. [R-UT] Мур, Гвен [D-WI] Морелл, Джозеф Д. [D-NY] Моултон, Сет [D-MA] Мрван, Фрэнк Дж. [D-IN] Маллин , Маркуэйн [R-OK] Мерфи, Грегори [R-NC] Мерфи, Стефани Н. [D-FL] Надлер, Джеррольд [D-NY] Наполитано, Грейс Ф. [D-CA] Нил, Ричард Э. [D -MA] Негус, Джо [D-CO] Нельс, Трой Э. [R-TX] Ньюхаус, Дэн [R-WA] Ньюман, Мари [D-IL] Норкросс, Дональд [D-NJ] Норман, Ральф [R -SC] Нортон, Элеонора Холмс [D-DC] Нуньес, Девин [R-CA] О’Халлеран, Том [D-AZ] Обернолте, Джей [R-CA] Окасио-Кортес, Александрия [D-NY] Омар, Ильхан [D-MN] Оуэнс, Берджесс [R-UT] Палаццо, Стивен М.[R-MS] Паллоне, Фрэнк-младший [D-NJ] Палмер, Гэри Дж. [R-AL] Панетта, Джимми [D-CA] Паппас, Крис [D-NH] Паскрелл, Билл-младший [D -NJ] Пейн, Дональд М.-младший [D-NJ] Пелоси, Нэнси [D-CA] Пенс, Грег [R-IN] Перлмуттер, Эд [D-CO] Перри, Скотт [R-PA] Питерс, Скотт Х. [D-CA] Пфлюгер, Август [R-TX] Филлипс, Дин [D-MN] Пингри, Челли [D-ME] Пласкетт, Стейси Э.
[R-WV] Миллер, Мэри Э. [R-IL] Миллер-Микс, Марианнетт [R-IA] Муленаар, Джон Р. [R-MI] Муни, Александр X. [R-WV] Мур, Барри [R -AL] Мур, Блейк Д. [R-UT] Мур, Гвен [D-WI] Морелл, Джозеф Д. [D-NY] Моултон, Сет [D-MA] Мрван, Фрэнк Дж. [D-IN] Маллин , Маркуэйн [R-OK] Мерфи, Грегори [R-NC] Мерфи, Стефани Н. [D-FL] Надлер, Джеррольд [D-NY] Наполитано, Грейс Ф. [D-CA] Нил, Ричард Э. [D -MA] Негус, Джо [D-CO] Нельс, Трой Э. [R-TX] Ньюхаус, Дэн [R-WA] Ньюман, Мари [D-IL] Норкросс, Дональд [D-NJ] Норман, Ральф [R -SC] Нортон, Элеонора Холмс [D-DC] Нуньес, Девин [R-CA] О’Халлеран, Том [D-AZ] Обернолте, Джей [R-CA] Окасио-Кортес, Александрия [D-NY] Омар, Ильхан [D-MN] Оуэнс, Берджесс [R-UT] Палаццо, Стивен М.[R-MS] Паллоне, Фрэнк-младший [D-NJ] Палмер, Гэри Дж. [R-AL] Панетта, Джимми [D-CA] Паппас, Крис [D-NH] Паскрелл, Билл-младший [D -NJ] Пейн, Дональд М.-младший [D-NJ] Пелоси, Нэнси [D-CA] Пенс, Грег [R-IN] Перлмуттер, Эд [D-CO] Перри, Скотт [R-PA] Питерс, Скотт Х. [D-CA] Пфлюгер, Август [R-TX] Филлипс, Дин [D-MN] Пингри, Челли [D-ME] Пласкетт, Стейси Э. [D-VI] Покан, Марк [D-WI] Портер, Кэти [D-CA] Поузи, Билл [R-FL] Прессли, Аянна [D-MA] Прайс, Дэвид Э. [D-NC] Куигли, Майк [D-IL] Радеваген, Аумуа Амата Коулман [R- А.С.] Раскин, Джейми [D-MD] Рид, Том [R-NY] Решенталер, Гай [R-PA] Райс, Кэтлин М.[D-NY]Райс, Том [R-SC]Ричмонд, Седрик Л. [D-LA]Роджерс, Гарольд [R-KY]Роджерс, Майк Д. [R-AL]Роуз, Джон В. [R-TN ] Розендейл-старший, Мэтью М. [R-MT] Росс, Дебора К. [D-NC] Роузер, Дэвид [R-NC] Рой, Чип [R-TX] Ройбал-Аллард, Люсиль [D-CA] Руис , Рауль [D-CA] Рупперсбергер, CA Dutch [D-MD] Раш, Бобби Л. [D-IL] Резерфорд, Джон Х. [R-FL] Райан, Тим [D-OH] Саблан, Грегорио Килили Камачо [ D-MP] Салазар, Мария Эльвира [R-FL] Сан-Николас, Майкл FQ [D-GU] Санчес, Линда Т. [D-CA] Сарбанес, Джон П. [D-MD] Скализ, Стив [R-LA ] Скэнлон, Мэри Гей [D-PA] Шаковски, Дженис Д.[D-IL]Шифф, Адам Б. [D-CA]Шнайдер, Брэдли Скотт [D-IL]Шредер, Курт [D-OR]Шриер, Ким [D-WA]Швайкерт, Дэвид [R-AZ]Скотт, Остин [R-GA] Скотт, Дэвид [D-GA] Скотт, Роберт С. «Бобби» [D-VA] Сешнс, Пит [R-TX] Сьюэлл, Терри А.
[D-VI] Покан, Марк [D-WI] Портер, Кэти [D-CA] Поузи, Билл [R-FL] Прессли, Аянна [D-MA] Прайс, Дэвид Э. [D-NC] Куигли, Майк [D-IL] Радеваген, Аумуа Амата Коулман [R- А.С.] Раскин, Джейми [D-MD] Рид, Том [R-NY] Решенталер, Гай [R-PA] Райс, Кэтлин М.[D-NY]Райс, Том [R-SC]Ричмонд, Седрик Л. [D-LA]Роджерс, Гарольд [R-KY]Роджерс, Майк Д. [R-AL]Роуз, Джон В. [R-TN ] Розендейл-старший, Мэтью М. [R-MT] Росс, Дебора К. [D-NC] Роузер, Дэвид [R-NC] Рой, Чип [R-TX] Ройбал-Аллард, Люсиль [D-CA] Руис , Рауль [D-CA] Рупперсбергер, CA Dutch [D-MD] Раш, Бобби Л. [D-IL] Резерфорд, Джон Х. [R-FL] Райан, Тим [D-OH] Саблан, Грегорио Килили Камачо [ D-MP] Салазар, Мария Эльвира [R-FL] Сан-Николас, Майкл FQ [D-GU] Санчес, Линда Т. [D-CA] Сарбанес, Джон П. [D-MD] Скализ, Стив [R-LA ] Скэнлон, Мэри Гей [D-PA] Шаковски, Дженис Д.[D-IL]Шифф, Адам Б. [D-CA]Шнайдер, Брэдли Скотт [D-IL]Шредер, Курт [D-OR]Шриер, Ким [D-WA]Швайкерт, Дэвид [R-AZ]Скотт, Остин [R-GA] Скотт, Дэвид [D-GA] Скотт, Роберт С. «Бобби» [D-VA] Сешнс, Пит [R-TX] Сьюэлл, Терри А. [D-AL] Шерман, Брэд [D -CA] Шерил, Мики [D-NJ] Симпсон, Майкл К. [R-ID] Сирс, Альбио [D-NJ] Слоткин, Элисса [D-MI] Смит, Адам [D-WA] Смит, Адриан [R -NE] Смит, Кристофер Х. [R-NJ] Смит, Джейсон [R-MO] Смакер, Ллойд [R-PA] Сото, Даррен [D-FL] Спанбергер, Эбигейл Дэвис [D-VA] Спартц, Виктория [ R-IN] Спейер, Джеки [D-CA] Стэнсбери, Мелани Энн [D-NM] Стэнтон, Грег [D-AZ] Стаубер, Пит [R-MN] Стил, Мишель [R-CA] Стефаник, Элиз М.[R-NY] Стейл, Брайан [R-WI] Стейбе, В. Грегори [R-FL] Стивенс, Хейли М. [D-MI] Стюарт, Крис [R-UT] Стиверс, Стив [R-OH] Стрикленд , Мэрилин [D-WA] Суоцци, Томас Р. [D-NY] Суолвелл, Эрик [D-CA] Такано, Марк [D-CA] Тейлор, Ван [R-TX] Тенни, Клаудия [R-NY] Томпсон , Бенни Г. [D-MS] Томпсон, Гленн [R-PA] Томпсон, Майк [D-CA] Тиффани, Томас П. [R-WI] Тиммонс, Уильям Р. IV [R-SC] Титус, Дина [ D-NV] Тлаиб, Рашида [D-MI] Тонко, Пол [D-NY] Торрес, Норма Дж. [D-CA] Торрес, Ричи [D-NY] Трэхан, Лори [D-MA] Троун, Дэвид Дж. .[D-MD] Тернер, Майкл Р. [R-OH] Андервуд, Лорен [D-IL] Аптон, Фред [R-MI] Валадао, Дэвид Г.
[D-AL] Шерман, Брэд [D -CA] Шерил, Мики [D-NJ] Симпсон, Майкл К. [R-ID] Сирс, Альбио [D-NJ] Слоткин, Элисса [D-MI] Смит, Адам [D-WA] Смит, Адриан [R -NE] Смит, Кристофер Х. [R-NJ] Смит, Джейсон [R-MO] Смакер, Ллойд [R-PA] Сото, Даррен [D-FL] Спанбергер, Эбигейл Дэвис [D-VA] Спартц, Виктория [ R-IN] Спейер, Джеки [D-CA] Стэнсбери, Мелани Энн [D-NM] Стэнтон, Грег [D-AZ] Стаубер, Пит [R-MN] Стил, Мишель [R-CA] Стефаник, Элиз М.[R-NY] Стейл, Брайан [R-WI] Стейбе, В. Грегори [R-FL] Стивенс, Хейли М. [D-MI] Стюарт, Крис [R-UT] Стиверс, Стив [R-OH] Стрикленд , Мэрилин [D-WA] Суоцци, Томас Р. [D-NY] Суолвелл, Эрик [D-CA] Такано, Марк [D-CA] Тейлор, Ван [R-TX] Тенни, Клаудия [R-NY] Томпсон , Бенни Г. [D-MS] Томпсон, Гленн [R-PA] Томпсон, Майк [D-CA] Тиффани, Томас П. [R-WI] Тиммонс, Уильям Р. IV [R-SC] Титус, Дина [ D-NV] Тлаиб, Рашида [D-MI] Тонко, Пол [D-NY] Торрес, Норма Дж. [D-CA] Торрес, Ричи [D-NY] Трэхан, Лори [D-MA] Троун, Дэвид Дж. .[D-MD] Тернер, Майкл Р. [R-OH] Андервуд, Лорен [D-IL] Аптон, Фред [R-MI] Валадао, Дэвид Г. [R-CA] Ван Дрю, Джефферсон [R-NJ] Ван Дайн, Бет [R-TX] Варгас, Хуан [D-CA] Визи, Марк А. [D-TX] Вела, Филемон [D-TX] Веласкес, Нидия М. [D-NY] Вагнер, Энн [R -MO] Уолберг, Тим [R-MI] Валорски, Джеки [R-IN] Вальц, Майкл [R-FL] Вассерман Шульц, Дебби [D-FL] Уотерс, Максин [D-CA] Уотсон Коулман, Бонни [D -NJ] Вебер, Рэнди К., старший [R-TX] Вебстер, Дэниел [R-FL] Уэлч, Питер [D-VT] Венструп, Брэд Р. [R-OH] Вестерман, Брюс [R-AR] Уэкстон, Дженнифер [D-VA] Уайлд, Сьюзен [D-PA] Уильямс, Никема [D-GA] Уильямс, Роджер [R-TX] Уилсон, Фредерика С.[D-FL] Уилсон, Джо [R-SC] Виттман, Роберт Дж. [R-VA] Вомак, Стив [R-AR] Райт, Рон [R-TX] Ярмут, Джон А. [D-KY] Янг , Дон [R-AK] Зелдин, Ли М. [R-NY] Любой член Сената Болдуин, Тэмми [D-WI] Баррассо, Джон [R-WY] Беннет, Майкл Ф. [D-CO] Блэкберн, Марша [ R-TN] Блюменталь, Ричард [D-CT] Блант, Рой [R-MO] Букер, Кори А. [D-NJ] Бузман, Джон [R-AR] Браун, Майк [R-IN] Браун, Шеррод [ D-OH] Берр, Ричард [R-NC] Кантвелл, Мария [D-WA] Капито, Шелли Мур [R-WV] Кардин, Бенджамин Л.
[R-CA] Ван Дрю, Джефферсон [R-NJ] Ван Дайн, Бет [R-TX] Варгас, Хуан [D-CA] Визи, Марк А. [D-TX] Вела, Филемон [D-TX] Веласкес, Нидия М. [D-NY] Вагнер, Энн [R -MO] Уолберг, Тим [R-MI] Валорски, Джеки [R-IN] Вальц, Майкл [R-FL] Вассерман Шульц, Дебби [D-FL] Уотерс, Максин [D-CA] Уотсон Коулман, Бонни [D -NJ] Вебер, Рэнди К., старший [R-TX] Вебстер, Дэниел [R-FL] Уэлч, Питер [D-VT] Венструп, Брэд Р. [R-OH] Вестерман, Брюс [R-AR] Уэкстон, Дженнифер [D-VA] Уайлд, Сьюзен [D-PA] Уильямс, Никема [D-GA] Уильямс, Роджер [R-TX] Уилсон, Фредерика С.[D-FL] Уилсон, Джо [R-SC] Виттман, Роберт Дж. [R-VA] Вомак, Стив [R-AR] Райт, Рон [R-TX] Ярмут, Джон А. [D-KY] Янг , Дон [R-AK] Зелдин, Ли М. [R-NY] Любой член Сената Болдуин, Тэмми [D-WI] Баррассо, Джон [R-WY] Беннет, Майкл Ф. [D-CO] Блэкберн, Марша [ R-TN] Блюменталь, Ричард [D-CT] Блант, Рой [R-MO] Букер, Кори А. [D-NJ] Бузман, Джон [R-AR] Браун, Майк [R-IN] Браун, Шеррод [ D-OH] Берр, Ричард [R-NC] Кантвелл, Мария [D-WA] Капито, Шелли Мур [R-WV] Кардин, Бенджамин Л. [D-MD] Карпер, Томас Р. [D-DE] Кейси , Роберт П., младший [D-PA] Кэссиди, Билл [R-LA] Коллинз, Сьюзен М. [R-ME] Кунс, Кристофер А. [D-DE] Корнин, Джон [R-TX] Кортес Масто, Кэтрин [D -NV] Коттон, Том [R-AR] Крамер, Кевин [R-ND] Крапо, Майк [R-ID] Круз, Тед [R-TX] Дэйнс, Стив [R-MT] Дакворт, Тэмми [D-IL] ] Дурбин, Ричард Дж. [D-IL] Эрнст, Джони [R-IA] Файнштейн, Дайэнн [D-CA] Фишер, Деб [R-NE] Гиллибранд, Кирстен Э. [D-NY] Грэм, Линдси [R -SC] Грассли, Чак [R-IA] Хагерти, Билл [R-TN] Харрис, Камала Д. [D-CA] Хассан, Маргарет Вуд [D-NH] Хоули, Джош [R-MO] Генрих, Мартин [ D-NM] Хикенлупер, Джон У.[D-CO] Хироно, Мэйзи К. [D-HI] Хувен, Джон [R-ND] Хайд-Смит, Синди [R-MS] Инхоф, Джеймс М. [R-OK] Джонсон, Рон [R-WI ] Кейн, Тим [D-VA] Келли, Марк [D-AZ] Кеннеди, Джон [R-LA] Кинг, Ангус С.-младший [I-ME] Клобучар, Эми [D-MN] Лэнкфорд, Джеймс [ R-OK] Лихи, Патрик Дж. [D-VT] Ли, Майк [R-UT] Леффлер, Келли [R-GA] Лухан, Бен Рэй [D-NM] Ламмис, Синтия М. [R-WY] Манчин , Джо, III [D-WV] Марки, Эдвард Дж.
[D-MD] Карпер, Томас Р. [D-DE] Кейси , Роберт П., младший [D-PA] Кэссиди, Билл [R-LA] Коллинз, Сьюзен М. [R-ME] Кунс, Кристофер А. [D-DE] Корнин, Джон [R-TX] Кортес Масто, Кэтрин [D -NV] Коттон, Том [R-AR] Крамер, Кевин [R-ND] Крапо, Майк [R-ID] Круз, Тед [R-TX] Дэйнс, Стив [R-MT] Дакворт, Тэмми [D-IL] ] Дурбин, Ричард Дж. [D-IL] Эрнст, Джони [R-IA] Файнштейн, Дайэнн [D-CA] Фишер, Деб [R-NE] Гиллибранд, Кирстен Э. [D-NY] Грэм, Линдси [R -SC] Грассли, Чак [R-IA] Хагерти, Билл [R-TN] Харрис, Камала Д. [D-CA] Хассан, Маргарет Вуд [D-NH] Хоули, Джош [R-MO] Генрих, Мартин [ D-NM] Хикенлупер, Джон У.[D-CO] Хироно, Мэйзи К. [D-HI] Хувен, Джон [R-ND] Хайд-Смит, Синди [R-MS] Инхоф, Джеймс М. [R-OK] Джонсон, Рон [R-WI ] Кейн, Тим [D-VA] Келли, Марк [D-AZ] Кеннеди, Джон [R-LA] Кинг, Ангус С.-младший [I-ME] Клобучар, Эми [D-MN] Лэнкфорд, Джеймс [ R-OK] Лихи, Патрик Дж. [D-VT] Ли, Майк [R-UT] Леффлер, Келли [R-GA] Лухан, Бен Рэй [D-NM] Ламмис, Синтия М. [R-WY] Манчин , Джо, III [D-WV] Марки, Эдвард Дж. [D-MA] Маршалл, Роджер [R-KS] МакКоннелл, Митч [R-KY] Менендес, Роберт [D-NJ] Меркли, Джефф [D-OR ] Моран, Джерри [R-KS] Мурковски, Лиза [R-AK] Мерфи, Кристофер [D-CT] Мюррей, Пэтти [D-WA] Оссофф, Джон [D-GA] Падилья, Алекс [D-CA] Пол , Рэнд [R-KY] Питерс, Гэри С.[D-MI] Портман, Роб [R-OH] Рид, Джек [D-RI] Риш, Джеймс Э. [R-ID] Ромни, Митт [R-UT] Розен, Джеки [D-NV] Раундс, Майк [R-SD] Рубио, Марко [R-FL] Сандерс, Бернард [I-VT] Сассе, Бен [R-NE] Шац, Брайан [D-HI] Шумер, Чарльз Э. [D-NY] Скотт, Рик [R-FL] Скотт, Тим [R-SC] Шахин, Жанна [D-NH] Шелби, Ричард С. [R-AL] Синема, Кирстен [D-AZ] Смит, Тина [D-MN] Стабеноу, Дебби [D-MI] Салливан, Дэн [R-AK] Тестер, Джон [D-MT] Тьюн, Джон [R-SD] Тиллис, Томас [R-NC] Туми, Патрик [R-PA] Тубервиль, Томми [R -AL] Ван Холлен, Крис [D-MD] Уорнер, Марк Р.[D-VA] Уорнок, Рафаэль Г. [D-GA] Уоррен, Элизабет [D-MA] Уайтхаус, Шелдон [D-RI] Уикер, Роджер Ф. [R-MS] Уайден, Рон [D-OR] Янг , Тодд [R-IN]
[D-MA] Маршалл, Роджер [R-KS] МакКоннелл, Митч [R-KY] Менендес, Роберт [D-NJ] Меркли, Джефф [D-OR ] Моран, Джерри [R-KS] Мурковски, Лиза [R-AK] Мерфи, Кристофер [D-CT] Мюррей, Пэтти [D-WA] Оссофф, Джон [D-GA] Падилья, Алекс [D-CA] Пол , Рэнд [R-KY] Питерс, Гэри С.[D-MI] Портман, Роб [R-OH] Рид, Джек [D-RI] Риш, Джеймс Э. [R-ID] Ромни, Митт [R-UT] Розен, Джеки [D-NV] Раундс, Майк [R-SD] Рубио, Марко [R-FL] Сандерс, Бернард [I-VT] Сассе, Бен [R-NE] Шац, Брайан [D-HI] Шумер, Чарльз Э. [D-NY] Скотт, Рик [R-FL] Скотт, Тим [R-SC] Шахин, Жанна [D-NH] Шелби, Ричард С. [R-AL] Синема, Кирстен [D-AZ] Смит, Тина [D-MN] Стабеноу, Дебби [D-MI] Салливан, Дэн [R-AK] Тестер, Джон [D-MT] Тьюн, Джон [R-SD] Тиллис, Томас [R-NC] Туми, Патрик [R-PA] Тубервиль, Томми [R -AL] Ван Холлен, Крис [D-MD] Уорнер, Марк Р.[D-VA] Уорнок, Рафаэль Г. [D-GA] Уоррен, Элизабет [D-MA] Уайтхаус, Шелдон [D-RI] Уикер, Роджер Ф. [R-MS] Уайден, Рон [D-OR] Янг , Тодд [R-IN]